ОСНОВЫ БИОХИМИИ ЛЕНИНДЖЕРА - ТОМ 1. ОСНОВЫ БИОХИМИИ СТРОЕНИЕ И КАТАЛИЗ - 2011
ЧАСТЬ I. СТРОЕНИЕ И КАТАЛИЗ
Чем крепче забор — тем дружней соседи.
Роберт Фрост, «Починка стены» в сб. «На север от Бостона», 1914
11. БИОЛОГИЧЕСКИЕ МЕМБРАНЫ И ТРАНСПОРТ
Первая клетка, вероятно, возникла, когда сформировалась мембрана, отделяющая небольшой объем водного раствора от остального мира. Мембраны образуют внешние границы клеток и регулируют движение молекул через эти границы (рис. 11-1). В клетках эукариот они делят внутреннее пространство на отдельные компартменты, чтобы обособить процессы и компоненты. Они организуют сложные последовательности реакций и являются главными как для запасания биологической энергии, так и для формирования межклеточных контактов. Биологические функции мембран проистекают из их замечательных физических свойств. Мембраны пластичны, способны к самогерметизации, избирательно проницаемы для полярных веществ. Благодаря пластичности мембран форма клеток может изменяться при клеточном росте и движении (например, при амебоидном движении). Обладая способностью к разрыву и замыканию, две мембраны могут сливаться, как при экзоцитозе; единичный ограниченный мембраной компартмент без больших утечек содержимого может делиться на два отдельных компартмента, как при эндоцитозе и клеточном делении. Поскольку мембраны обладают избирательной проницаемостью, они удерживают некоторые вещества и ионы внутри клеток и внутри отдельных клеточных компартментов, не пропуская внутрь другие.
Рис. 11-1. Биологические мембраны. На поперечном срезе все клеточные мембраны имеют характерный трехслойный вид. После окрашивания эритроцита тетроксидом осмия его плазматическая мембрана выглядит в электронном микроскопе как трехслойная структура толщиной от 5 до 8 нм (50-80 А). Трехслойное изображение состоит из двух электроноплотных слоев, разделенных менее плотной центральной областью (осмий связывается с внутренней и внешней поверхностями мембраны).
Мембраны не просто пассивные барьеры. В них содержится множество белков, предназначенных для облегчения или катализа различных клеточных процессов. На клеточной поверхности переносчики перемещают специфические органические вещества и неорганические ионы через мембрану; рецепторы воспринимают внеклеточ-
ные сигналы и запускают молекулярные изменения в клетке; адгезивные молекулы удерживают вместе соседние клетки. Внутри клетки мембраны организуют клеточные процессы, такие как синтез липидов и некоторых белков, и перенос энергии в митохондриях и хлоропластах. Поскольку мембраны состоят только из двух молекулярных слоев, они очень тонкие — практически двумерные. Межмолекулярные столкновения в этом двумерном пространстве гораздо более вероятны, так что эффективность ферментативных процессов в мембране чрезвычайно увеличивается.
В этой главе мы прежде всего опишем состав клеточных мембран и их химическое строение — молекулярные структуры, которые лежат в основе их биологических функций. Затем рассмотрим замечательные динамические свойства мембран, в которых липиды и белки взаимно перемещаются. Обсудим динамическую роль мембранных белков при клеточной адгезии, эндоцитозе и слиянии мембран, сопровождающем секрецию нейромедиаторов. Потом мы обратимся к опосредованному белками перемещению веществ через мембраны при помощи переносчиков и ионных каналов. В последующих главах мы обсудим роль мембран в передаче сигналов (гл. 12 и 23), переносе энергии (гл. 19), синтезе липидов (гл. 21) и синтезе белков (гл. 27).
11.1. Состав и строение мембран
Для понимания функционирования мембран важное значение имеет изучение их состава - какие молекулы встречаются во всех мембранах, а какие уникальны для мембран со специфическими функциями. Поэтому, прежде чем перейти к структуре и функциям мембран, рассмотрим молекулярный состав биологических мембран: белки и полярные липиды, которые составляют основную часть мембран, и углеводы (гликопротеины и гликолипиды).
Каждый тип мембран содержит характерные липиды и белки
Относительное содержание белков и липидов меняется в зависимости от типа мембран (табл. 11-1), отражая различие биологических функций. Например, некоторые нейроны имеют миелиновую оболочку — протяженную плазматическую мембрану, которая многократно окружает клетку и действует как пассивный электрический изолятор. Миелиновая оболочка в основном состоит из липидов, в то время как плазматические мембраны бактерий и мембраны митохондрий и хлоропластов, в которых сосредоточены многие ферментативные процессы, содержат больше (по массе) белков, чем липидов.
Таблица 11-1. Состав плазматических мембран у различных организмов
Компоненты (% по массе) Стерин Другие липиды Белки |
Фосфолипиды |
Стерины |
|
Миелиновая оболочка нейронов человека |
30 30 19 |
холестерин |
галактолипиды, плазмалогены |
Печень мыши |
45 27 25 |
холестерин |
- |
Листья кукурузы |
47 26 7 |
ситостерин |
галактолипиды |
Дрожжи |
52 7 4 |
эргостерин |
триацилглицерины, эфиры стеринов |
р. Paramecium (инфузория) |
56 40 4 |
стигмастерин |
- |
Е. coli |
75 25 0 |
- |
- |
Примечание: суммарный состав нс достигает 100 %, поскольку кроме белков, фосфолипидов и стеринов имеются и другие вещества. Например, растения содержат большие количества гликолипидом.
Для изучения состава мембраны первой задачей является ее изоляция. Когда клетки подвергаются механическому измельчению, их плазматическая мембрана разрывается и фрагментируется, при этом высвобождаются цитоплазматические компоненты и мембраносвязанные органеллы — митохондрии, хлоропласты, лизосомы и ядра. Фрагменты плазматической мембраны и интактные органеллы можно выделить путем центрифугирования (рис. 1-8 и пример 2-1 на с. 87).
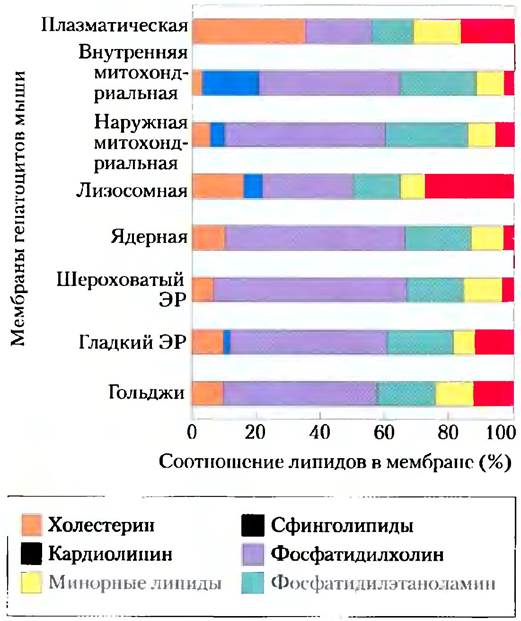
В клетках, несомненно, есть механизмы контроля типов и количества мембранных липидов на пути от их синтеза до доставки конкретных липидов в определенные органеллы. Каждое царство, каждый вид, каждая ткань или тип клеток, а также органеллы каждого типа клеток обладают характерным набором мембранных липидов. Например, плазматические мембраны обогащены холестерином и не содержат сколько-нибудь заметных количеств кардиолипина (рис. 11-2); во внутренних мембранах митохондрий гепатоцитов распределение обратное: очень мало холестерина и много кардиолипина. За редким исключением, мы можем говорить об адаптивной роли разных комбинаций мембранных липидов; в других случаях функциональное значение этих комбинаций пока неизвестно.
Рис. 11-2. Липидный состав плазматических мембран и мембран органелл гепатоцитов крысы. Функциональная специализация каждого типа мембраны отражается в уникальном составе липидов. Холестерин в значительных количествах присутствует в плазматических мембранах, но обнаруживается в следовых количествах в мембранах митохондриальных. Кардиолипин — существенный компонент внутренней митохондриальной, но не плазматической мембраны. В большинстве мембран фосфатидилсерин, фосфатидилинозит и фосфатидилглицерин — минорные компоненты (желтый цвет), но они выполняют важнейшие функции, например, фосфатидилинозит и его производные участвуют в передаче гормональных сигналов. Сфинголипиды, фосфатидилхолин и фосфатидилэтаноламин присутствуют в большинстве мембран, но в разных соотношениях. Гликолипидов много в мембранах хлоропластов растений, а в тканях животных они практически отсутствуют.
Белковый состав мембран изменяется даже в более широких пределах, чем липидный, отражая их функциональную специализацию. Кроме того, некоторые мембранные белки ковалентно связаны с олигосахаридами. Например, 60% (по массе) молекул гликофорина — гликопротеина мембраны эритроцитов — состоит из сложных олигосахаридных единиц, ковалентно присоединенных к специфическим аминокислотным остаткам. Чаще всего сахара присоединяются по остаткам Ser, Thr и Asn (рис. 7-29). Остатки сахаров поверхностных гликопротеинов влияют на фолдинг белков, а также на их стабильность и внутриклеточную локализацию. Они играют существенную роль в специфическом связывании лигандов с гликопротеиновыми поверхностными рецепторами (рис. 7-35).
Некоторые мембранные белки ковалентно связаны с липидами (одним или более), служащими гидрофобными якорями, которые, как мы увидим, удерживают белки в мембране.
Все биологические мембраны обладают рядом фундаментальных свойств
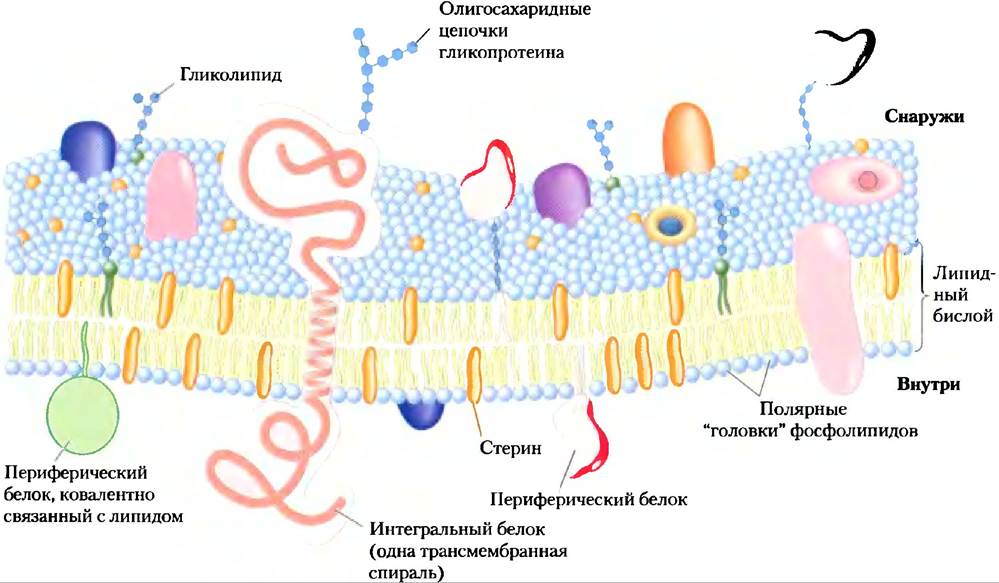
Мембраны непроницаемы для большинства полярных или заряженных веществ, но проницаемы для неполярных соединений; толщина мембран от 5 до 8 нм (50-80 А); на поперечном срезе в электронном микроскопе они выглядят трехслойными (рис. 11-1). По данным электронной микроскопии, химического анализа состава, а также физических методов исследования проницаемости мембран и движения отдельных молекул липидов и белков внутри мембраны создана жидкостно-мозаичная модель биологических мембран (рис. 11-3). Фосфолипиды образуют бислой, где неполярные части молекул обращены внутрь бислоя, а их полярные «головки» обращены наружу и взаимодействуют с водной фазой с обеих сторон мембраны. Белки включаются в этот бислойный пласт, удерживаясь между мембранными липидами и гидрофобными доменами белков благодаря своим гидрофобным взаимодействиям. Некоторые белки «высовываются» только с одной стороны мембраны, в других есть домены, экспонированные с обеих сторон. Ориентация белков в бислое асимметрична, что придает мембране «двусторонность»: белковые домены на одной стороне бислоя отличаются от доменов на другой стороне, что отражает и функциональную асимметрию. Отдельные липидные и белковые единицы в мембране формируют постоянно изменяющуюся жидкую мозаичную структуру (в отличие от мозаики керамической плитки, уложенной на цемент). Мембранная мозаика — жидкая, так как большинство взаимодействий между ее компонентами нековалентные; липиды и белки способны свободно двигаться вдоль плоскости мембраны.
Рис. 11-3. Жидкостно-мозаичная модель структуры мембраны. Жирнокислотные цепочки внутри мембраны формируют гидрофобную область. Интегральные белки плавают в этом липидном «море» благодаря своим гидрофобным свойствам, обусловленным неполярными боковыми аминокислотными цепями. И белки, и липиды могут свободно двигаться вдоль плоскости бислоя, но движение с одной стороны бислоя на другую ограничено. Углеводные части, связанные с некоторыми белками и липидами плазматической мембраны, находятся на внешней поверхности мембраны.
Теперь рассмотрим некоторые особенности жидкостно-кристаллической модели более детально и приведем экспериментальные доказательства в пользу основной идеи предложенной модели, однако с необходимыми усовершенствованиями.
Липидный бислой — основной структурный элемент биомембран
Глицерофосфолипиды, сфинголипиды и стерины практически нерастворимы в воде. При смешивании с водой они самопроизвольно образуют микроскопические липидные агрегаты (кластеры) в виде обособленной фазы, при этом их гидрофобные части находятся в контакте друг с другом, а гидрофильные группы взаимодействуют с окружающей водной средой. Вспомним, что при объединении липидов в кластеры уменьшается площадь гидрофобной поверхности, обращенной в сторону воды, и таким образом минимизируется число молекул из упорядоченной воды на поверхности раздела липид-вода (рис. 2-7), что приводит к увеличению энтропии. Благодаря гидрофобным взаимодействиям между молекулами липидов создается термодинамическая движущая сила, обеспечивающая образование и сохранение этих кластеров.
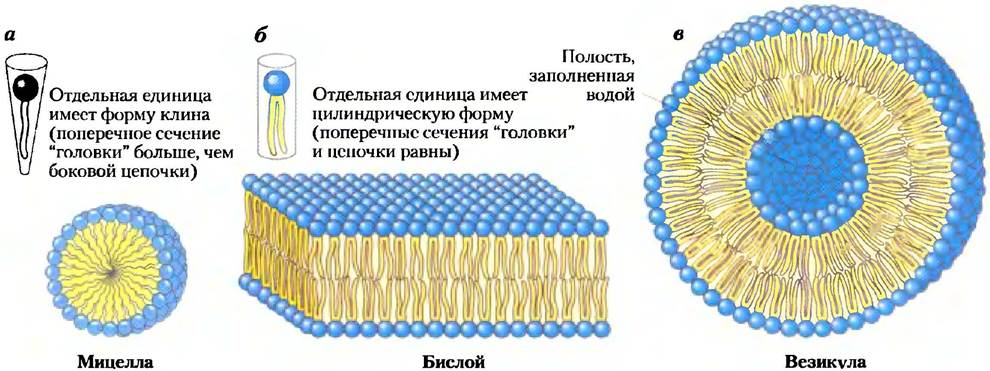
При смешивании липидов с водой в зависимости от условий и природы липидов могут образоваться три типа липидных кластеров (рис. 11-4). Мицеллы имеют сферическую форму и не содержат ничего, кроме амфифильных молекул (от нескольких десятков до нескольких тысяч). Эти молекулы расположены таким образом, что их гидрофобные области агрегированы внутри, куда вода не попадает, а их гидрофильные «головки» — к поверхности мембраны и контактируют с водной средой. Мицеллообразование облегчается, когда площадь поперечного сечения «головки» больше, чем площадь сечения боковой ацильной цепи (цепей), как в свободных жирных кислотах, лизофосфолипидах (фосфолипидах без одной жирной кислоты) и детергентах, таких как додецилсульфат натрия (SDS; с. 136).
Второй тип липидного агрегата в воде — это бислой, в котором два липидных монослоя образуют двумерный лист. Легче всего образование бислоя происходит, когда полплощади поперечных сечений «головки» и ацильной цепи (цепей) близки, как в глицерофосфолипидах и сфинголипидах. Гидрофобные области каждого монослоя взаимодействуют друг с другом, не касаясь воды. Гидрофильные «головки» взаимодействуют с водой на обеих поверхностях бислоя. Поскольку на краях гидрофобные области (рис. 11-4, б) имеют кратковременные контакты с водой, бислойный пласт относительно нестабилен и спонтанно сворачивается, накладываясь сам на себя, образуя полую сферу — везикулу (пузырек) (рис. 11-4, в). При образовании везикул бислои теряют свои концевые гидрофобные области, достигая максимальной стабильности в водном окружении. Внутри этих бислойных везикул заключена вода в виде отдельного водного компартмента. Вероятно, предшественники первых живых клеток напоминали липидные пузырьки, их водное содержимое было отделено от остального мира гидрофобной облочкой.
Рис. 11-4. Агрегаты из амфифильных липидов, образующиеся в воде, а) В мицеллах гидрофобные цепи жирных кислот собраны в центре сферы. Внутри гидрофобной области практически нет воды. б) В открытом бислое все ацильные боковые цепочки, кроме тех, которые расположены на краях пласта, защищены от взаимодействия с водой, в) Когда двумерный бислой сворачивается, он образует замкнутый бислой, трехмерный полый пузырек (везикулу), в полости которого находится вода.
Биологические мембраны состоят из липидных бислоев толщиной 3 нм (30 А). Углеводородная часть мембраны, составленная из групп —СН2- и -СН3 жирных кислот, неполярна и похожа в этом отношении надекан, и везикулы, сформированные в лаборатории из чистых липидов (липосомы), почти непроницаемы для полярных веществ, как и биологические мембраны (хотя последние проницаемы для веществ при наличии специфических переносчиков).
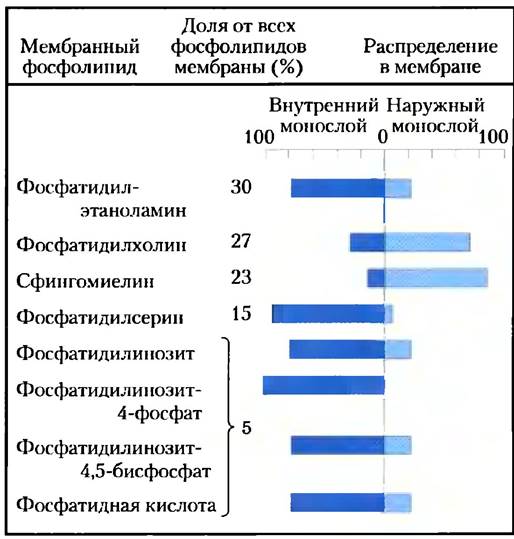
Липиды плазматической мембраны асимметрично распределены между двумя монослоями, хотя в отличие от мембранных белков эта асимметрия не абсолютна. В плазматической мембране эритроцита, например, холинсодержащие липиды (фосфатидилхолин и сфингомиелин) обычно находятся во внешнем (экзоплазматическом) монослое (рис. 11-5), в то время как фосфатидилсерин, фосфатидилэтаноламин и фосфатидилинозиты гораздо более часто встречаются во внутреннем (цитоплазматическом) монослое. Изменения в распределении липидов между монослоями плазматической мембраны приводят к серьезным последствиям. Например, только когда фосфатидил серин в плазматической мембране переходит в наружный монослой, тромбоцит способен исполнить свою роль в образовании кровяного сгустка. Для многих других типов клеток наличие фосфатидилсерина на внешней поверхности мембраны служит «меткой» для разрушения клетки путем программируемой клеточной смерти.
Рис. 11-5. Асимметричное распределение фосфолипидов между внутренним и наружным монослоями плазматической мембраны эритроцита. Распределение специфического фосфолипида определяется при обработке интактной клетки фосфолипазой С, которая не может подействовать на липиды внутреннего монослоя, но удаляет «головки» липидов внешнего монослоя. Содержание высвобожденных «головок» является мерой количества каждого липида в наружном монослое.
Три типа мембранных белков различаются расположением в мембране
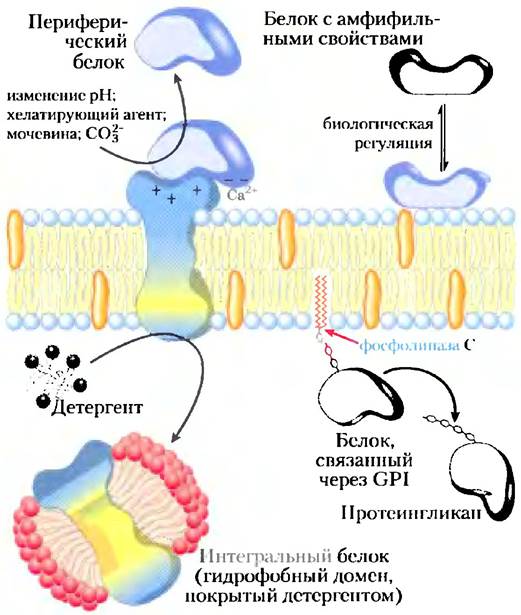
Интегральные мембранные белкиочень прочно связаны с мембраной, они удаляются только с помощью агентов, которые нарушают гидрофобные взаимодействия, — детергентов, органических растворителей или денатурирующих веществ (рис. 11-6). Периферические мембранные белки связываются с мембраной посредством электростатических взаимодействий или образования водородных связей с гидрофильными доменами интегральных белков и с полярными «головками» мембранных липидов. Они могут высвобождаться после относительно мягкой обработки, влияющей на электростатические взаимодействия или разрушающей водородные связи; для этого часто добавляют карбонат при высоких pH. Амфитропные белки могут как находиться в цитозоле, так и быть связанными с мембраной. Их сродство к мембранам в некоторых случаях определяется нековалентными взаимодействиями с мембранными белками или липидами, а в других случаях объясняется наличием одного или нескольких липидов, связанных с амфитропным белком ковалентной связью (см. рис. 11-14). Обычно обратимое связывание амфитропных белков с мембранами регулируется; например, фосфорилирование или связывание лиганда может привести к конформационным изменениям в белке, в результате которых открывается ранее недоступный участок связывания с мембраной.
Рис. 11-6. Периферические, интегральные и амфитропные белки. Мембранные белки можно разделить в соответствии с условиями, необходимыми для их высвобождения из мембраны. Большинство периферических белков высвобождаются при изменении pH или ионной силы, удаления Са2+-хелатирующими агентами или при добавлении мочевины или карбоната. Интегральные белки экстрагируются детергентами, нарушающими гидрофобные взаимодействия с липидным бислоем и образующими мицеллоподобные кластеры вокруг отдельных белковых молекул. Интегральные белки, ковалентно связанные с мембранными липидами, такими как гликозилфосфатидилинозит (GPI, рис. 11-14), можно выделить, обработав фосфолипазой С. Амфитропные белки иногда соединены с мембраной, а иногда нет; процесс связывания регулируется, например, с помощью обратимой реакции пальмитилирования.
Многие мембранные белки пронизывают липидный бислой
Топологию мембранного белка (его локализацию относительно липидного бислоя) можно определить с помощью реагентов, которые взаимодействуют с боковыми цепями белка, но нс могут пересекать мембраны. Это, например, полярные химические реагенты, которые взаимодействуют с первичными аминами остатков лизина, или ферменты, подобные трипсину, которые расщепляют белки, но не могут проходить через мембрану. Удобный объект для таких исследований — эритроциты человека, так как они не содержат мембраносвязанных органелл, а единственная их мембрана — плазматическая. Если мембранный белок в интактном эритроците реагирует с агентом, не проходящим через мембрану, этот белок должен иметь по крайней мере один домен, расположенный на внешней (внеклеточной) стороне мембраны. Трипсин расщепляет внеклеточные домены, но не влияет на домены, скрытые внутри бислоя или расположенные исключительно на внутренней поверхности, если только плазматическая мембрана не разрушается специально, чтобы сделать эти домены доступными для фермента.
Эксперименты с такими топологически специфическими реагентами показывают, что гликопротеин эритроцитов гликофорин пронизывает плазматическую мембрану. Его N-концевой домен (несущий углеводные цепи) расположен на внешней поверхности и расщепляется трипсином. Карбоксильный конец выступает из мембраны внутри клетки, где он нс может реагировать с непроникающими реагентами. И N-, и С-концевые домены содержат много полярных или заряженных аминокислотных остатков и поэтому полностью гидрофильны. Однако сегмент в центре белка (остатки 75-93) содержит много гидрофобных аминокислотных участков, и это приводит нас к заключению, что гликофорин имеет трансмембранный сегмент, расположенный так, как показано на рис. 11-7.
Рис. 11-7. Гликофорин в эритроците пронизывает бислой. Один гидрофильный домен, содержащий все сахарные остатки, находится на внешней поверхности, а другой гидрофильный домен «высовывается» из внутренней поверхности мембраны. Каждый красный шестиугольник изображает тетрасахарид, содержащий две молекулы Neu5Ac (сиаловая кислота). Gal и GalNAc, связанные через кислород с остатком серина (Ser) или треонина (Thr); синий шестиугольник — олигосахаридная цепь, связанная через атом N с остатком Asn. В реальности у олигосахаридных единиц относительный размер больше. Сегмент из 19 гидрофобных остатков (75-93) образуют α-спираль, которая пересекает мембранный бислой (рис. 11-11, а). Сегмент из остатков аминокислот 64-74 содержит несколько гидрофобных остатков и, вероятно, погружен в мембрану с внешней стороны, как показано на рисунке.
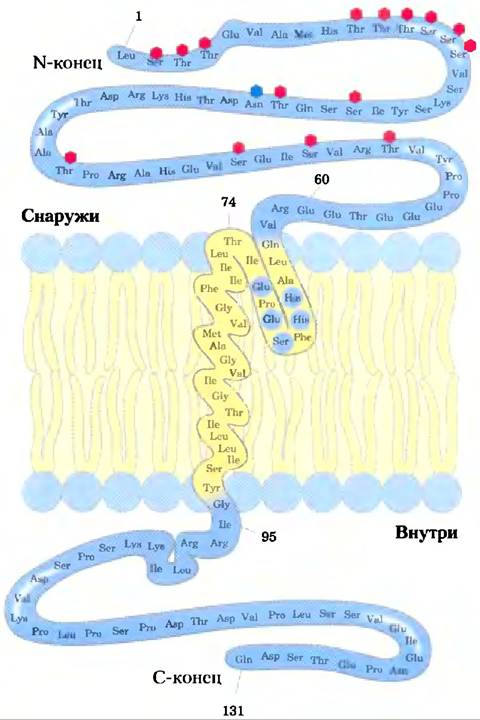
Экспериментально выявлено, что гликофорин в мембране расположен асимметрично: 14-конец всегда направлен наружу. Аналогично для других мембранных белков показано, что каждый белок обладает своей специфической ориентацией в бислое: один домен трансмембранного белка всегда направлен наружу, другой — внутрь. Кроме того, гликопротеины плазматической мембраны всегда располагаются таким образом, что их сахарные остатки находятся на внешней поверхности клетки. Как мы увидим, асимметричное расположение мембранных белков приводит к функциональной асимметрии. Если рассмотреть строение ионного насоса, например, выясняется, что все его молекулы имеют одну и ту же ориентацию в мембране и, следовательно, работают в одном направлении.
Интегральные белки взаимодействуют своими гидрофобными доменами с липидами и благодаря этому удерживаются в мембране
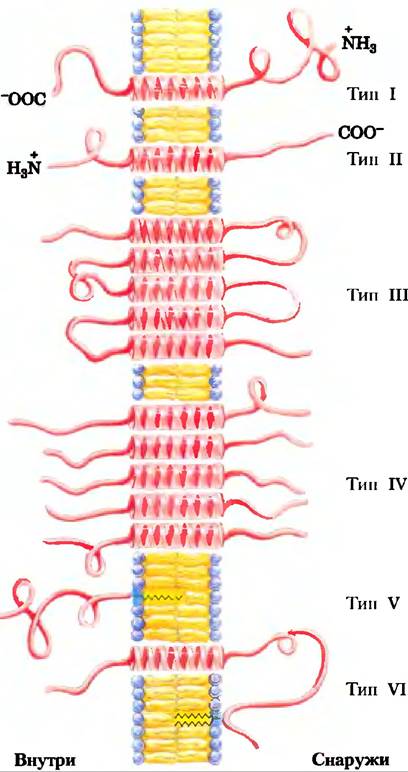
Прочное соединение интегральных белков с мембранами достигается благодаря гидрофобным взаимодействиям между мембранными липидами и гидрофобными доменами белка. Некоторые белки содержат единственную гидрофобную последовательность в середине (как гликофорин), на N- или С-конце. У других белков есть множественные гидрофобные последовательности, каждая из которых, если она имеет α-спиральную конформацию, достаточно протяженна, чтобы пронизывать липидный бислой (рис. 11-8).
Рис. 11-8. Интегральные мембранные белки. Известные белки плазматической мембраны по пространственным отношениям белковых доменов и липидного бислоя можно отнести к шести типам. Типы I и II содержат только одну трансмембранную α-спиралъ; N-концевой домен находится снаружи в белке типа I и внутри — в белке типа II. Белки типа III имеют множественные трансмембранные спирали в одном полипептиде. В белках типа IV трансмембранные домены нескольких разных полипептидов соединяются с образованием канала через мембрану. Белки типа V удерживаются бислоем прежде всего с помощью ковалентно связанных липидов (рис. 11-14), белки типа VI содержат и трансмембранные спирали, и липидные (GРI) якори.
На этом рисунке и на протяжении всей книги мы представляем трансмембранные белковые сегменты в их наиболее вероятных конформациях — как α-спирали из шести-семи витков. Иногда эти спирали показаны просто в виде цилиндров. Поскольку относительно мало мембранных белковых структур было расшифровано методом РGА, изображение внемембранных доменов условно и необязательно соответствует истинному масштабу.
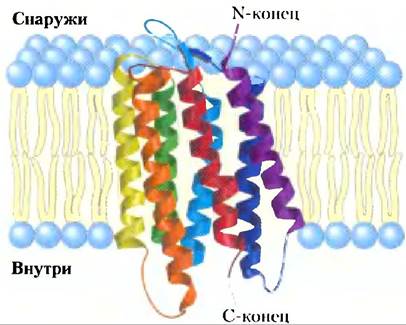
Один из наиболее изученных белков, пронизывающих мембрану, бактериородопсин содержит семь сильногидрофобных внутренних последовательностей и пересекает липидный бислой семь раз. Бактериородопсин — это запускающийся светом протонный насос, плотно упакованный в виде упорядоченных массивов в пурпурной мембране бактерии Halobacterium salinarium. С помощью рентгеновской кристаллографии была установлена структура с семью α-спиральными сегментами (каждый пронизывает липидный бислой), связанными неспиральными петлями на внешней и внутренней сторонах мембраны (рис. 11-9). В аминокислотной последовательности бактериродоисина идентифицируются семь сегментов, состоящих каждый примерно из 20 гидрофобных остатков, т. е. длина каждого сегмента как раз достаточна для образования пересекающей бислой α-спирали. Семь спиралей вместе образуют кластер и ориентированы почти перпендикулярно плоскости бислоя, обеспечивая трансмембранный путь для движения протонов. Как мы увидим в гл. 12, этот набор из семи гидрофобных пронизывающих мембрану спиралей является общим мотивом для мембранных белков, участвующих в восприятии сигналов. Гидрофобные взаимодействия между неполярными аминокислотами и жирноацильными группами мембранных липидов прочно заякоривают белок в мембране.
Рис. 11-9. Бактериородопсин — пронизывающий мембрану белок (РDВ ID 2АТ9). Единственная полипептидная цепь складывается в семь гидрофобных α-спиралей, каждая из которых пересекает липидный бислой почти перпендикулярно к плоскости мембраны. Семь трансмембранных спиралей объединяются в кластер, и пространство вокруг них и между ними заполняется ацильными цепями мембранных липидов. Светопоглощающий пигмент ретиналь (рис. 10-21) погружен глубоко в мембрану и контактирует с несколькими спиральными сегментами (не показаны). Степень окраски спиралей соответствует их гидрофильности (см. рис. 11-11, б).
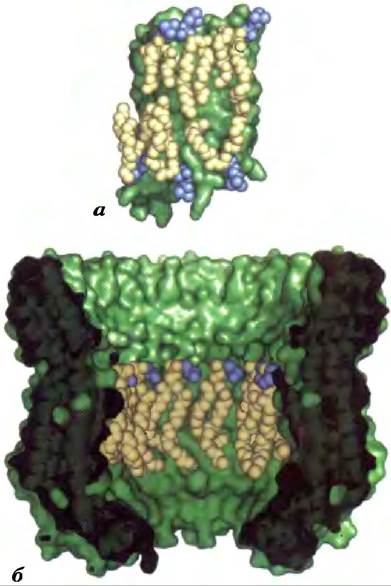
Исследованные с помощью методов кристаллографии мембранные белки (таким образом было установлено их молекулярное строение) часто содержат молекулы фосфолипидов; считается, что они обнаруживаются в кристаллах, потому что находятся в исходных мембранах. Многие из молекул фосфолипидов лежат на поверхности белков, их «головки» взаимодействуют с полярными остатками аминокислот на границе мембраны и водной среды с внутренней и внешней сторон, а боковые цепи ассоциированы с неполярными остатками. Эти кольцевые липиды образуют бислойную раковину (кольцо) вокруг белка, ориентированную приблизительно так, как и положено фосфолипидам в бислое (рис. 11-10). Другие фосфолипиды были обнаружены на границе мономеров в мультисубъединичных мембранных белках, где они формировали «уплотнение из смазки». Кроме того, фосфолипиды могут быть глубоко внедрены в мембранные белки, обычно их «головки» находятся под уровнем мембраны. Например, у сукцинатдегидрогеназы (комплекс II, встречается в митохондриях; см. рис. 19-10) есть несколько глубоко утопленных молекул фосфолипидов.
Рис. 11-10. Липидные кольца, связанные с двумя интегральными мембранными белками, а) Кристаллическая структура аквапорина овцы (трансмембранный водный канал; PDBID 2В60), состоящего из фосфолипидной оболочки, в которой «головки» молекул (синие) располагаются на внешней и внутренней поверхностях мембраны, а гидрофобные цепочки (бежевые) тесно связаны с поверхностью белка. Липид образует жировой слой вокруг белка, изображенный в виде поверхности зеленого цвета. б) Кристаллическая структура интегрального белкового комплекса Na+-АТРазы типа V из Enterococcus hirae (PDB ID 2BL2), состоящего из 10 идентичных субъединиц, каждая из которых содержит четыре трансмембранные спирали, окружающие центральную полость, в которой расположен фосфатидилглицерин (ФГ). На данном рисунке пять субъединиц удалены, чтобы дать возможность читателю увидеть молекулы фосфатидилглицерина, связанные с каждой субъединицей.
Трехмерную структуру мембранного белка можно предсказать по его последовательности
Обычно определение трехмерной структуры (топологии) мембранного белка — гораздо более сложная задача, чем определение аминокислотной последовательности секвенированием белка или его гена. Известны тысячи аминокислотных последовательностей мембранных белков, однако с помощью РСА или ЯМР было установлено гораздо меньше трехмерных структур. Присутствие непрерывных последовательностей из более чем 20 гидрофобных остатков в мембранном белке принято считать доказательством того, что эти последовательности погружены в липидный бислой, где они работают как гидрофобные якори или образуя трансмембранные каналы. Практически каждый интегральный белок имеет по крайней мере одну такую последовательность. Если так же рассуждать в случае геномных последовательностей, то окажется, что у многих видов от 20% до 30% всех белков — интегральные мембранные белки.
Какие можно сделать предсказания относительно вторичной структуры интегральных белков, которые пересекают мембрану? α-Спиральная последовательность из 20-25 остатков имеет длину как раз достаточную, чтобы пронизывать всю толщину (30 А) липидного бислоя (напомним, что длина α-спирали составляет 1,5 А (0,15 нм) на один аминокислотный остаток). Полипептидная цепь, окруженная липидами, не имеет возможности образовать водородные связи с молекулами воды и стремится перейти к структуре α-спирали или β-слоя, где количество водородных связей максимально. Если боковые цепи всех аминокислот в спирали неполярны, гидрофобные взаимодействия с окружающими липидами будут еще сильнее стабилизировать спираль.
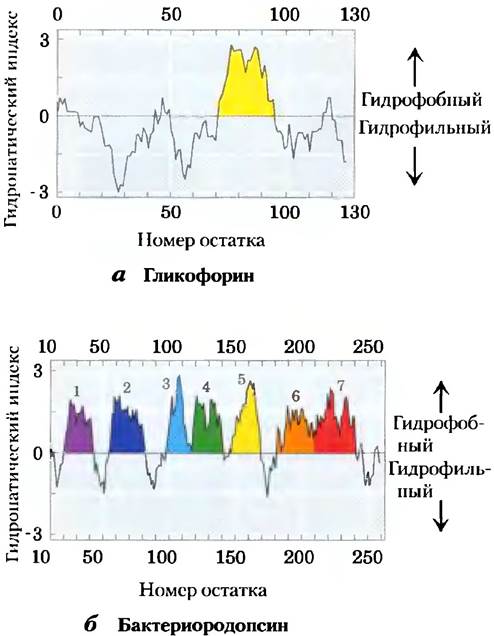
Существует несколько простых методов анализа аминокислотных последовательностей, дающих возможность достаточно точно предсказать вторичную структуру трансмембранных белков. Относительную полярность каждой аминокислоты определяют экспериментально, измеряя изменение свободной энергии, сопровождающее перемещение аминокислотной боковой цепи из гидрофобного растворителя в воду. Энергетика этого процесса, которую можно выразить через индекс гидрофобности(«гидропатический» индекс) для данной аминокислотной последовательности (табл. 3-1), колеблется в пределах от очень экзергонической для заряженных или полярных остатков вплоть до очень эндергонической для аминокислот с ароматическими или алифатическими углеводородными боковыми цепями. Общая гидрофобность последовательности аминокислот оценивается путем суммирования свободных энергий переноса аминокислотных остатков в последовательности. Чтобы проверить полипептидную последовательность на наличие сегментов, пронизывающих мембрану, исследователь рассчитывает индекс гидрофобности данной последовательности определенного размера — от 7 до 20 остатков, называемой окном. Для окон из семи остатков, например, от 1 до 7, от 2 до 8, от 3 до 9 и т. д« индексы гидрофобности наносят на график, как показано на рис. 11-11 (в каждом окне выбрана середина окна, например остаток 4 для интервала с 1 по 7). Область, содержащая больше чем 20 остатков с высоким индексом гидрофобности, считается трансмембранным сегментом. Когда последовательности мембранных белков с известной трехмерной структурой сканируют таким образом, обнаруживается достаточно хорошее соответствие между предсказанным и известным трансмембранными сегментами. С помощью анализа гидрофобности можно предсказать наличие единственной гидрофобной спирали в гликофорине (рис. 11-11, а) и семи трансмембранных сегментов в бактериородопсине (рис. 11-11, б), что соответствует результатам экспериментов.
Рис. 11-11. Профили гидрофобности. Индекс гидрофобности (табл. 3-1) отложен напротив номеров остатков двух интегральных белков. Индекс гидрофобности для каждого аминокислотного остатка в последовательности определенной длины (окна) используется для расчета средней гидрофобности остатков этого окна. По оси абсцисс — номер остатка в середине окна, а) Гликофорин эритроцитов человека содержит единственную гидрофобную последовательность между остатками 75 и 93 (желтый цвет); ср. с рис. 11-7. б) По данным исследований различными методами, у бактериородопсина семь трансмембранных спиралей (рис. 11-9) и семь гидрофобных областей. Заметим, однако, что в области сегментов 6 и 7 профиль гидрофобности допускает двоякое толкование. Исследования подтвердили, что в этой области два трансмембранных сегмента.
На основе аминокислотных последовательностей и кривых гидрофобности предполагается, что многие транспортные белки, описанные в этой главе, содержат множественные погруженные в мембрану спиральные участки, иными словами, они принадлежат к интегральным белкам типа III или IV (рис. 11-8). Если результаты согласуются с химическими исследованиями локализации белка (как описано для гликофорина и бактериородопсина), вероятность того, что гидрофобные области представляют собой именно пересекающие мембрану домены, существенно возрастает.
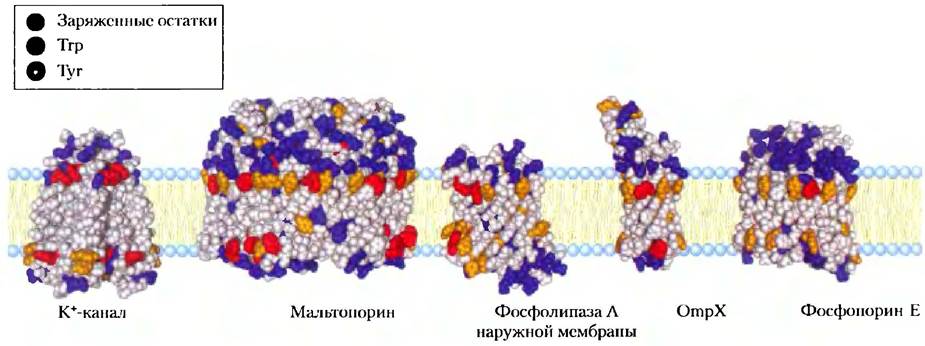
Еще одним замечательным качеством многих трансмембранных белков, структура которых расшифрована, является присутствие остатков Туr и Тrр на поверхности раздела между липидом и водой (рис. 11-12). Боковые цепи этих остатков служат, по всей видимости, якорями на поверхности раздела, способными одновременно взаимодействовать с центральной липидной фазой и водными фазами на каждой стороне мембраны. Еще одно общее наблюдение о расположении аминокислотных остатков относительно двойного липидного слоя состоит в следующем: положительно заряженные остатки Lys, His и Arg в мембранных белках чаще всего встречаются на цитоплазматической стороне мембраны (правило «положительные остатки — внутри»).
Рис. 11-12. Остатки Туr и Тrр мембранных белков, образующих кластеры на поверхности раздела вода-липид. По данным кристаллографических исследований была рассчитана структура этих пяти интегральных мембранных белков. К+-канал (PDB ID 1BL8) из Streptomyces lividans (рис. 11-48); мальтопорин (PDB ID 1AF6), фосфолипаза А внешней мембраны (PDB ID 1QD5), OmpX (PDB ID 1QJ9) и фосфопорин Е (PDB ID 1РНO) — белки наружной мембраны E. coli. Остатки Туr (оранжевый цвет) и Тrр (красный) обнаруживаются преимущественно в тех местах, где неполярная область ацильных цепей входит в контакт с областью полярных головок. Заряженные остатки (Lys, Arg, Glu, Asp) окрашены в голубой цвет; они обнаруживаются почти исключительно в водных фазах.
Не все интегральные мембранные белки составлены из трансмембранных α-спиралей. Другим структурным мотивом, общим для мембранных белков, является β-цилиндр (рис. 4-17, г), в котором 20 или больше трансмембранных сегментов формируют β-слои, выстраивающиеся в виде цилиндра (рис. 11-13). Те же самые факторы, которые благоприятствуют образованию α-спирали в гидрофобном окружении липидного бислоя, стабилизируют и β-цилиндры. Когда нет молекул воды для образования водородной связи с карбонильным кислородом или азотом пептидной связи, наиболее стабильную конформацию обеспечивает максимально возможное количество водородных связей между цепочками. В плоских ß-слоях водородными связями связаны не все атомы, которые способны к их образованию, и β-слои, как правило, не обнаружены в мембранных белках; β-цилиндры допускают образование всех возможных водородных связей и характерны для мембранных белков. Порины — белки, которые позволяют некоторым полярным веществам пересекать наружную мембрану грамотрицательных бактерий, таких как Е. coli, содержат многоцепочечные β-цилиндры, формирующие полярный трансмембранный коридор.
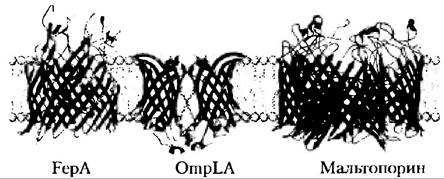
Рис. 11-13. Мембранные белки с β-цилиндрической (бочкообразной) структурой. В плоскости мембраны показаны три белка наружной мембраны E. coli. Белок FepA (PDB ID 1FEP), участвующий в обмене железа, содержит 22 пересекающие мембрану β-цепочки. Белок OmpLA (из PDB ID 1QD5), фосфолипаза, — β-цилиндр из 12 складок (в мембране существует в виде димера). Мальтопорин (из PDB ID 1MAL), переносчик мальтозы, — тример, каждый мономер которого состоит из 16 β-складок.
Полипетиды чаще принимают β-конформацию, чем α-спираль; для пересечения мембраны в β-конформации достаточно от семи до девяти остатков. Напомним, что в β-конформации вариабельные части отходят поочередно то вверх, то вниз от плоскости (рис. 4-6). В β-структурах мембранных белков каждый второй остаток в пронизывающем мембрану сегменте гидрофобный и взаимодействует с липидным бислоем; ароматические вариабельные части остатков обычно находятся на границе раздела липид-белок. Другие остатки могут быть или не быть гидрофильными. В случае белков с β-структурными мотивами для предсказания трансмембранных сегментов профили гидрофобности непригодны. Однако поскольку база данных, содержащая такие мотивы, увеличивается, предсказания трансмембранных β-конформаций, основанные на аминокислотной последовательности, становятся осуществимыми. Например, на основании анализа последовательности было правильно предсказано, что ряд мембранных белков грамотрицательных бактерий (рис. 11-13) содержат β-цилиндры.
Ковалентно связанные липиды удерживают некоторые мембранные белки
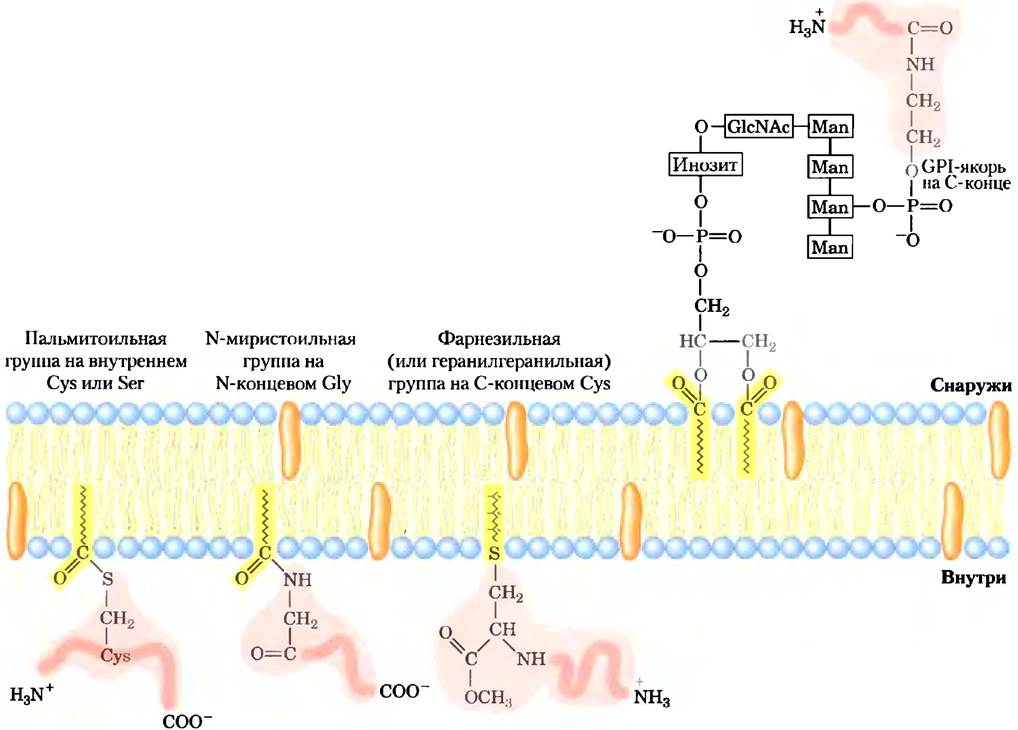
Некоторые мембранные белки ковалентно связываются с липидами (с одним или более) нескольких типов: с длинноцепочечными жирными кислотами, изопреноидами, стеринами или гликозилированными производными фосфатидил- инозита (GРI; рис. 11-14). Эти связанные липиды служат гидрофобным якорем, включенным в липидный бислой, благодаря которому белок удерживается на поверхности мембраны. Сила гидрофобного взаимодействия между бислоем и отдельной углеводородной цепочкой, связанной с белком, может оказаться недостаточной для того, чтобы надежно заякорить белок, но у многих белков есть не один связывающий липидный участок. Другие взаимодействия, такие как ионное притяжение между положительно заряженными остатками Lys в белке и отрицательно заряженными «головками» липидов, вероятно, тоже вносят свой вклад в стабилизацию соединения. Связь этих липид-связанных белков с мембраной, конечно, слабее, чем у интегральных мембранных белков, по крайней мере, в некоторых случаях она легко рвется. Однако при обработке карбонатами щелочных металлов GPI- связанные белки не высвобождаются, таким образом, их можно считать интегральными.
Помимо простого заякоривания белка на мембране, связанный липид может играть специфическую роль. В плазматической мембране белки с GPI-якорями находятся исключительно на внешней поверхности и образуют скопления в определенных участках, как мы увидим ниже (с. 543-546), в то время как другие связанные с липидами белки (с присоединенными фарнезильными или геранилгеранильными группами; рис. 11-14) присутствуют только на внутренней поверхности. В вытянутых эпителиальных клетках (таких как эпителиальные клетки кишечника; см. рис. 11-44), в которых апикальная и базальная поверхности выполняют разные функции, GPI-связанные белки направляются именно к апикальной поверхности. Присоединение специфического липида к вновь синтезированному мембранному белку, таким образом, управляет, локализуя этот белок.
Рис. 11-14. Мембранные белки, связанные с липидами. Ковалентно связанные липиды заякоривают мембранные белки в липидном бислое. Пальмитоильная группа связана тиоэфирной связью с остатком Суs; N-миристоильная группа обычно присоединяется к Gly на N-конце; фарнезильная и геранилгеранильная группы, связанные с остатками Суsна С-конце, являются изопреноидами, содержащими соответственно 15 и 20 атомов углерода. Эти три липид-белковых ансамбля находятся только на внутренней стороне плазматической мембраны. Гликозилфосфати- дилинозитные (GPI) якори представляют собой производные фосфатидилинозита, в котором инозит несет короткий олигосахарид, ковалентно присоединенный к С-концевому остатку белка через фосфоэтаноламин. GPI-связанные белки всегда расположены на внеклеточной поверхности плазматической мембраны.
Краткое содержание раздела 11.1 Состав и строение мембран
■ Биологические мембраны определяют границы клеток, делят клетки на дискретные компартменты, организуют сложные последовательности реакций и действуют в процессах приема сигналов и превращений энергии.
■ Мембраны построены из липидов и белков в разных комбинациях, характерных для каждого вида, типа клеток и органелл. Жидкостно-мозаичная модель описывает свойства, общие для всех биологических мембран. Основной структурной единицей мембран является липидный бислой.
■ Периферические мембранные белки нежестко связаны с мембраной посредством электростатических взаимодействий и водородных связей или с помощью ковалентно присоединенных липидных якорных молекул. Интегральные белки прочно связываются с мембраной с помощью гидрофобных взаимодействий между липидным бислоем и неполярными радикалами остатков аминокислот, которые экспонированы на поверхности белковой молекулы. Амфифильные белки связываются с мембраной обратимым образом.
■ Многие мембранные белки пронизывают липидный бислой несколько раз, при этом гидрофобные последовательности примерно из 20 аминокислотных остатков образуют трансмембранные α-спирали. Для интегральных мембранных белков также характерны многоцепочечные β-цилиндры. Остатки Туr и Тrр трансмембранных белков, как правило, находятся на границе липид-вода.
■ Существует специфичность расположения включенных в мембрану липидов и белков на той или иной стороне мембраны. Таким образом, мембраны структурно и функционально асимметричны. Многие мембранные белки содержат ковалентно связанные олигосахариды. Гликопротеины плазматической мембраны всегда ориентированы так, что несущий углевод домен находится на внеклеточной поверхности мембраны.