ОСНОВЫ БИОХИМИИ ЛЕНИНДЖЕРА - ТОМ 1. ОСНОВЫ БИОХИМИИ СТРОЕНИЕ И КАТАЛИЗ - 2011
ЧАСТЬ I. СТРОЕНИЕ И КАТАЛИЗ
11. БИОЛОГИЧЕСКИЕ МЕМБРАНЫ И ТРАНСПОРТ
11.2. Динамика мембран
Одно из поразительных свойств всех биологических мембран — их пластичность, т. е. способность изменять форму без потери целостности и образования течи. Это обусловлено наличием нековалентных взаимодействий между липидами в бислое и возможностью движения отдельных молекул липидов, поскольку они связаны друг с другом нековалентно. Теперь обратимся к динамике мембран: движениям мембраны и промежуточным структурам, которые могут при этом возникать.
Ацильные группы внутри бислоя упорядочены в разной степени
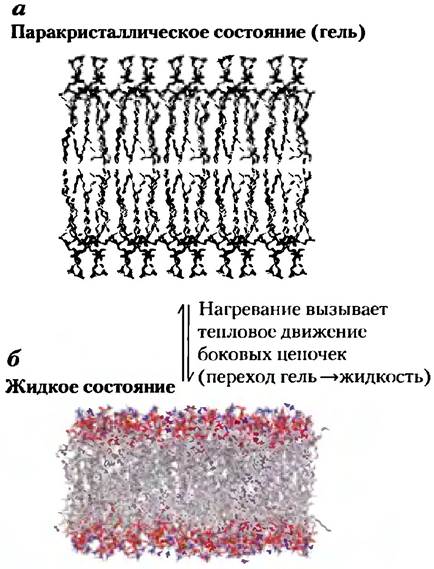
Хотя структура липидного бислоя весьма стабильна, отдельные молекулы фосфолипидов и стеринов обладают некоторой свободой движения (рис. 11-15). Структура и стабильность липидного бислоя зависят от присутствующих липидов и меняются при изменении температуры. При температурах ниже физиологических липиды в бислое образуют полутвердую фазу — гель, в котором все движения отдельных молекул липидов строго ограничены: бислой является паракристаллическим (рис. 11-15, а). При температурах выше физиологических углеводородные цепи жирных кислот находятся в постоянном движении, обусловленном вращением вокруг углерод- углеродных связей длинных ацильных боковых цепей. В этом жидко-разупорядоченном (или жидком) состоянии (рис. 11-15, в) внутренняя область бислоя напоминает море из находящихся в постоянном движении липидов. При промежуточных температурах липиды существуют в жидко-упорядоченном состоянии; здесь в меньшей степени проявляется тепловое движение в ацильных цепях липидного бислоя, но латеральное движение еще происходит. Эти различия в состоянии бислоя легко наблюдать в липосомах, сформированных из липидов одного типа, но биологические мембраны содержат много липидов с разнообразными ацильными цепями и поэтому не демонстрируют резких фазовых переходов при изменении температуры.
Рис. 11-15. Два состояния липидов бислоя, а) В паракристаллическом состоянии (фазе геля) полярные «головки» равномерно выстроены на поверхности, а ацильные цепи почти неподвижны и упакованы в регулярную геометрическую структуру, б) В жидко-неупорядоченном (жидком) состоянии ацильные цепи в большей степени подвержены тепловому движению и не имеют регулярной организации. Промежуточным между этими крайними состояниями является жидко-упорядоченное состояние, при котором отдельные фосфолипидные молекулы могут диффундировать латерально, но ацильные группы остаются вытянутыми и более или менее упорядоченными.
При физиологических температурах (20- 40 °С) длинноцепочечные насыщенные жирные кислоты (такие как 16:0 и 18:0) хорошо упаковываются в жидко-упорядоченную структуру, но изгибы в ненасыщенных жирных кислотах препятствуют такой упаковке (см. рис. 10-2), создавая благоприятные условия для жидконеупорядоченного состояния. Тот же эффект оказывают ацильные группы с более короткими цепочками. Содержание стеринов в мембране (которое существенно различается для разных организмов и органелл; табл. 11-1) — другой важный фактор, определяющий состояние липидов. Жесткая планарная структура стероидного ядра, внедренная между жирными ацильными боковыми цепями, уменьшает свободу вращательного движения соседних жирных ацильных цепей вокруг их углерод-углеродных связей, заставляя ацильные цени принять полностью вытянутую конформацию. Таким образом, присутствие стеринов уменьшает текучесть в центре бислоя, благоприятствуя гем самым жидко-упорядоченной фазе, и увеличивает толщину липидного монослоя (как описано ниже).
Таблица 11-2.
Содержание жир |
ных кислот, % * |
|||
10 °С |
20 °С |
30 °С |
40 °С |
|
Миристиновая кислота (14:0) |
4 |
4 |
1 |
8 |
Пальмитиновая кислота (16:0) |
18 |
25 |
29 |
18 |
Пальмитоленноная кислота (16:1) |
26 |
21 |
23 |
9 |
Олеиновая кислота (18:1) |
38 |
24 |
30 |
12 |
Гидроксимиристиновая кислота |
13 |
10 |
10 |
8 |
Относительное содержание**, насыщенные кислоты/ ненасыщенные кислоты |
2.9 |
2,0 |
1,6 |
0,38 |
* Точный жирнокислотный состав зависит нс только от температуры, но и от сталии роста и состава среды культивирования.
** Отношение суммарного содержания кислот (16:1) + (18:1) к суммарному содержанию кислот (14:0) + (16:0). Гидроксимиристиновая кислота не учитывалась.
При росте в разных условиях клетки регулируют свой липидный состав для достижения постоянной текучести мембраны. Например, бактерии, когда их культивируют при низкой температуре, синтезируют больше ненасыщенных жирных кислот и меньше насыщенных, чем при повышенных температурах (табл. 11-2). В результате такой регуляции состава липидов мембраны бактерий, культивированных как при высокой, так и при низкой температуре, имеют примерно одинаковую степень текучести.
Для движения липидов через бислой необходим катализ
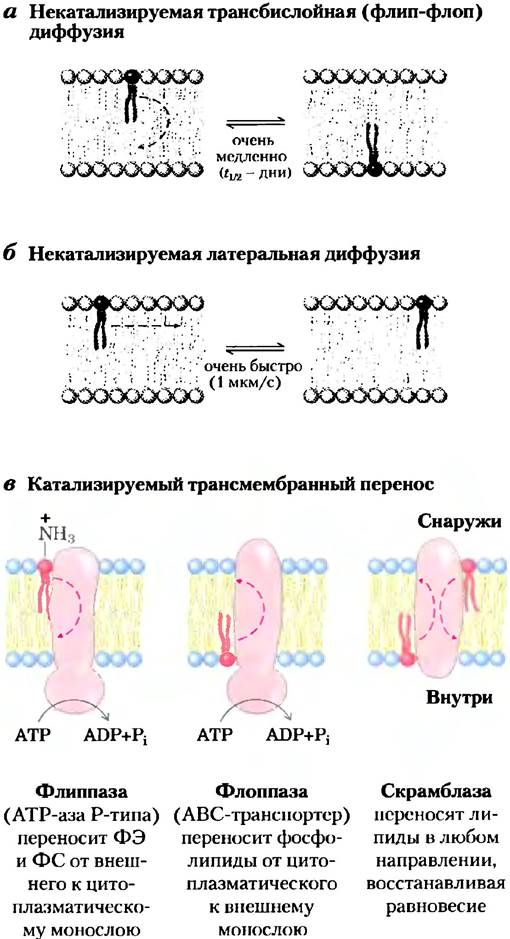
При физиологической температуре диффузия молекул липидов от одного монослоя к другому («флип-флоп»; рис. 11-16, а) в большинстве мембран либо вообще не случается, либо ее скорость очень низка, а вот латеральная диффузия в плоскости двойного слоя происходит очень быстро (рис. 11-16, б). Для движения через бислой необходимо, чтобы полярная или заряженная «головка» покинула свое водное окружение и перешла в гидрофобное внутреннее пространство бислоя, а этот процесс требует большого положительного изменения свободной энергии. Однако существуют ситуации, когда такое движение необходимо. Например, мембранные глицерофосфолипиды синтезируются на цитоплазматической поверхности ЭР, а сфинго- липиды синтезируются или модифицируются на люминальной поверхности. Перемещение липидов от места синтеза к месту назначения осуществляется путем флип-флоп-диффузии.
Существуют целые семейства белков, включая флиппазы, флоппазы и скрамблазы (рис. 11-16, в), которые облегчают перемещение липидов сквозь двойной слой, обеспечивая энергетически более выгодный и более быстрый путь, чем в отсутствие катализа. Сочетание асимметрии биосинтеза мембранных липидов, очень медленной некатализируемой флип-флоп-диффузии и наличия селективных энергозависимых переносчиков липидов приводит к асимметрии липидного состава в слоях мембраны (рис. 11-5). Кроме вклада в эту асимметрию состава энергозависимый транспорт липидов к одной стороне двойного слоя может оказывать важное влияние — путем создания более развитой поверхности на одной из сторон двойного слоя — на возникновение изгибов мембраны, необходимых для отделения везикул.
Рис. 11-16. Перемещение отдельных фосфолипидов в двойном липидном слое, а) Некатализируемое перемещение от одного монослоя к другому происходит очень медленно, в то время как латеральная диффузия (б) происходит быстро и не требует участия катализаторов. а) Три типа переносчиков фосфолипидов в плазматической мембране. Флиппазы переносят главным образом аминофосфолипиды (фосфатидилэтаноламин (ФЭ) и фосфатидилсерин (ФС)) от внешнего (экзоплазматического) к внутреннему (цитоплазматическому) монослою; этим представителям семейства АТРаз P-типа для проявления активности требуется АТР. Флоппазы переносят фосфолипиды от цитозольного монослоя к внешнему монослою; они являются членами семейства ABC-транспортеров и также используют АТР. Скрамблазы выравнивают фосфолипидный состав двух монослоев; они не используют АТР, но активируются в присутствии ионов кальция.
Флиппазы катализируют перенос аминофосфолипидов фосфатидилэтаноламина и фосфатидилсерина от внеклеточного монослоя к цитозольному монослою плазматической мембраны, внося свой вклад в асимметрию распределения фосфолипидов: фосфатидилэтаноламин и фосфатидилсерин располагаются в основном на цитозольной поверхности, а сфинголипиды и фосфатидилходлин — на внешней поверхности. Локализация фосфатидилсерина не на внешней поверхности мембраны имеет особый смысл: его появление на внешней поверхности мембраны запускает апоптоз (программируемую клеточную смерть, см. гл. 12) и поглощение макрофагами, несущими рецепторы фосфатидилсерина. Флиппазы действуют также в ЭР, где они перемещают вновь синтезированные фосфолипиды от места синтеза на цитозольной поверхности к люминальной поверхности. Для перемещения одной молекулы фосфолипида флиппазы затрачивают примерно одну молекулу АТР; по структуре и функциям эти белки родственны АТРазам P-типа (активные переносчики; см. с. 558). Флоппазы перемещают фосфолипиды плазматической мембраны от цитозольной к внеклеточной поверхности и, подобно флиппазам, являются ATP-зависимыми ферментами. Флоппазы — члены семейства АВС-транспортеров, описанного на с. 564, которые активно переносят гидрофобные субстраты через плазматическую мембрану наружу. Скрамблазы — это белки, которые переносят любые мембранные фосфолипиды через двойной липидный слой по градиенту концентрации (от поверхности с большей концентрацией к поверхности с меньшей концентрацией); их активность не зависит от присутствия АТР. Действие скрамблаз приводит к контролируемому перемешиванию состава липидных «головок» на обеих сторонах двойного слоя. Активность этих белков сильно возрастает при увеличении концентрации ионов кальция в цитозоле, которое может быть следствием активации клеток, их повреждения или апоптоза. Как отмечалось выше, появление фосфатидилсерина на внешней поверхности мембраны готовит клетку к апоптозу и поглощению макрофагами. Наконец, еще одну группу белков составляют транспортеры фосфатидилинозита, основная функция которых состоит в перемещении фосфатидилинозитов через двойной липидный слой; считается, что эти белки играют важную роль в передаче сигнала и мембранном транспорте.
Липиды и белки латерально диффундируют в бислое
Молекулы липидов могут участвовать в латеральной диффузии в мембране, меняясь местами с соседними молекулами липидов, т. е. совершать броуновское движение в пределах двойного слоя (рис. 11-16, б); такие перемещения могут быть очень быстрыми. Молекула в одном монослое липидного бислоя, например, внешнем монослое плазматической мембраны эритроцита, может диффундировать столь быстро, что «проплывает» эритроцит за секунды. Из-за быстрой латеральной диффузии внутри плоскости бислоя положение молекул случайно меняется в течение нескольких секунд.
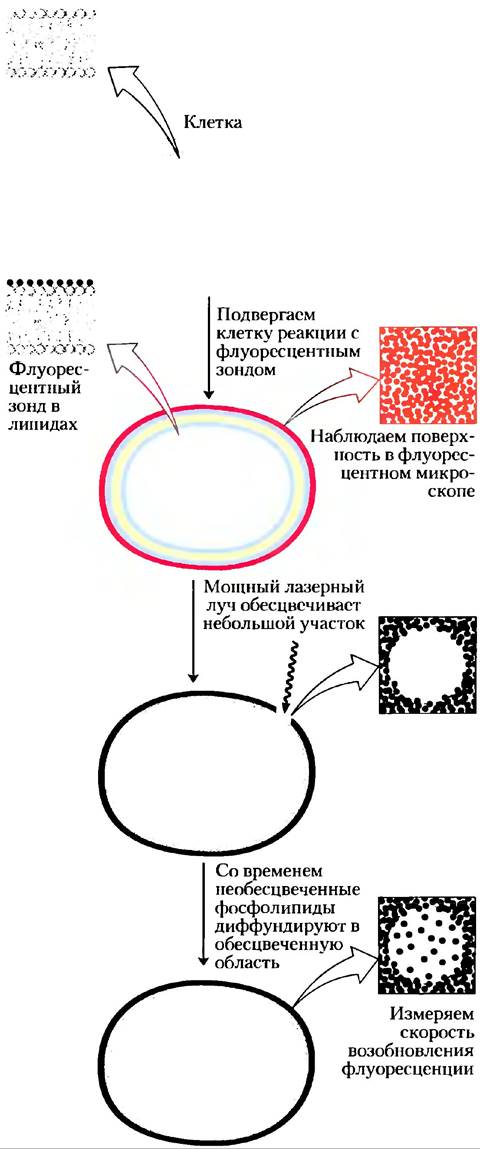
Латеральную диффузию можно продемонстрировать экспериментально посредством присоединения флуоресцентных зондов к «головкам» липидов и использования флуоресцентной спектроскопии, для отслеживания поведения зондов во времени (рис. 11-17). Но с одной из методик небольшая область клеточной поверхности (5 мкм2) с флуоресцентно меченными липидами обесцвечивается интенсивным лазерным облучением так, чтобы облученное пятно больше не флуоресцировало, при наблюдении в гораздо более слабом свете флуоресцентного микроскопа. Однако в течение миллисекунд наблюдаемый участок восстанавливает свою флуоресценцию, по мере того как «необесцвеченные» липидные молекулы диффундируют в область «обесцвеченного» пятна, а «обесцвеченные» — из него. Мерой скорости латеральной диффузии липидов является скорость возобновления флуоресценции после фотообесцвечивания (FRAP от англ. fluorescence recovery after photobleaching). При использовании метода FRAP было показано, что некоторые мембранные липиды диффундируют со скоростью до 1 мкм/с.
Рис. 11-17. Измерение скоростей латеральной диффузии липидов методом обесцвечивания флуоресценции (FRAP). Липиды в наружном монослое плазматической мембраны метят непроникающим сквозь мембрану флуоресцентным зондом (красный цвет), так что при наблюдении с помощью флуоресцентного микроскопа поверхность мембраны равномерно флуоресцирует. Маленькое пятно обесцвечивается при интенсивном облучении лазерным лучом (эта область не флуоресцирует). Со временем меченые липидные молекулы диффундируют в обесцвеченную область, и она снова становится флуоресцирующей. Из кинетики возобновления флуоресценции определяется коэффициент диффузии меченого липида. Обычно скорости диффузии липидов высокие; липид, движущийся с этой скоростью, может «обойти» Е. coli за 1 с. (Метод FRAP можно также использовать для измерения латеральной диффузии мембранных белков.)
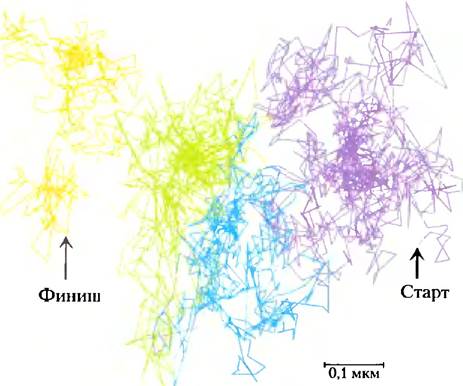
Другая методика отслеживания одиночной частицы позволяет наблюдать за движением одной-единственной молекулы в плазматической мембране за очень короткий промежуток времени. Результаты таких исследований подтверждают быструю латеральную диффузию внутри малой выделенной области поверхности клетки и показывают, что движение от одной такой области к соседней области затруднено; мембранные липиды ведут себя так, как если бы они были помещены в загон, огороженный забором, через который они могли бы время от времени перескакивать (рис. 11-18).
Рис. 11-18. Скачкообразная диффузия отдельных липидных молекул. С помощью флуоресцентного микроскопа сделана видеозапись движения отдельной флуоресцирующей липидной молекулы на поверхности клетки с разрешением 25 мкс (что эквивалентно 40 000 кадров в секунду). Показан трек молекулы, за которой наблюдали в течение 56 мс (всего 2250 кадров; путь начинается в фиолетовой области и продолжается через голубую, зеленую и оранжевую). Картина движения указывает на быструю диффузию внутри ограниченной области (около 250 нм в диаметре, она окрашена одним цветом) со случайными перескоками в прилежащую область. Напрашивается аналогия, что липиды словно огорожены «молекулярной изгородью», которую они время от времени перепрыгивают.
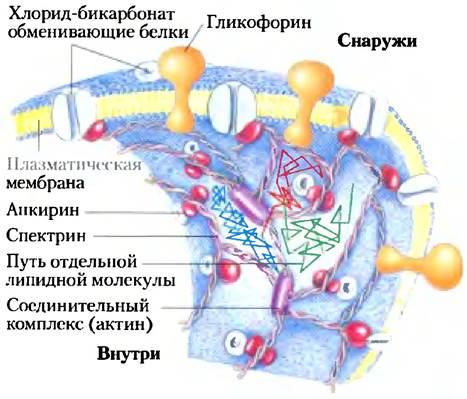
Многие мембранные белки, по-видимому, «плавают» в море липидов. Подобно мембранным липидам, эти белки могут латерально диффундировать в плоскости бислоя и находятся в постоянном движении, что подтверждается методом FRAP с флуоресцентно меченными поверхностными белками. Некоторые мембранные белки соединяются в пэтчи (от англ. patch — заплатка) — большие агрегаты на поверхности клетки или органеллы, в которых молекулы не могут двигаться друг относительно друга; например, ацетилхолиновые рецепторы образуют плотные пэтчи на плазматической мембране нейрона в области синапсов. Другие мембранные белки заякориваются на внутренних структурах, что предотвращает их свободную диффузию. В мембране эритроцита и гликофорин, и хлоридно-бикарбонатный обменник (с. 556) привязаны к спектрину, нитевидному белку цитоскелета (рис. 11-19). Одним из возможных объяснений представленной на рис. 11-18 картины латеральной диффузии липидных молекул является то, что мембранные белки, иммобилизованные посредством связывания со спектрином, служат «изгородью», которая определяет области относительно неограниченного движения липидов.
Рис. 11-19. Ограниченное движение хлоридно- бикарбонатного обменника и гликофорина в эритроцитах. Белки пронизывают мембрану и присоединяются к спектрину, белку цитоскелета, с помощью другого белка — анкирина, при этом ограничивается их способность к латеральному перемещению. Анкирин заякоривается в мембране ковалентно связанной пальмитоильной боковой цепочкой (рис. 11-14). Спектрин, длинный нитевидный белок, образует поперечную сшивку с соединительным комплексом, содержащим актин. Сеть поперечносшитых молекул спектрина, связанных с внутренней стороной плазматической мембраны, делает мембрану устойчивой к деформации. Эта сеть заякоренных мембранных белков служит «загоном» в эксперименте, показанном на рис. 11-18; движения липидов ограничены областями, содержащими связанные мембранные белки.
Сфинголипиды и холестерин объединяются в кластеры — мембранные рафты
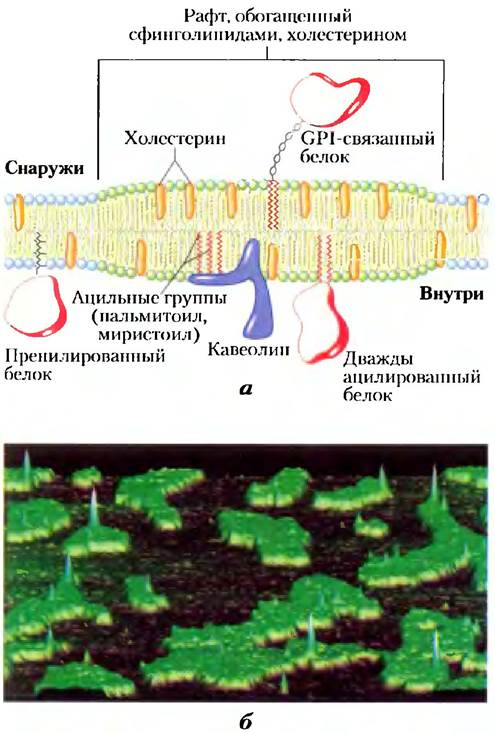
Мы видели, что диффузия мембранных липидов из одного монослоя в другой в случае, если она не катализируется, протекает очень медленно и что разные виды липидов плазматической мембраны распределены в этих монослоях асимметрично (рис. 11-5). Даже внутри одного монослоя распределение липидов небеспорядочно. Гликосфинголипиды (цереброзиды и ганглиозиды), содержащие обычно длинноцепочечные насыщенные жирные кислоты, образуют во внешнем монослое промежуточные кластеры, в которых практически отсутствуют глицерофосфолипиды, имеющие, как правило, одну ненасыщенную ацильную жирнокислотную группу и более короткую насыщенную жирнокислотную группу. Длинные насыщенные ацильные группы сфинголипидов могут образовывать более компактные, более стабильные связи с длинной кольцевой системой холестерина, чем более короткие, часто ненасыщенные цепи фосфолипидов. Состоящие из холестерина и сфинголипидов микродомены в наружном монослое плазматической мембраны, видимые с помощью атомно-силовой микроскопии (доп. 11-1), несколько толще и более упорядочены (менее жидкие), чем прилежащие микродомены, обогащенные фосфолипидами. Они с большим трудом растворяются неионными детергентами и ведут себя, как жидко-упорядоченные сфинголипидные плоты (рафты), дрейфующие в океане жидко-разупорядоченных фосфолипидов (рис. 11-20).
Рис. 11-20. Микродомены (рафты) в мембране, а) Стабильные ассоциаты сфинголипидов и холестерина во внешнем монослое образуют микродомен несколько большей толщины, чем другие области мембраны, обогащенный специфическими видами мембранных белков. В наружном монослое таких рафтов наиболее часто обнаруживаются GPI-связанные белки, а белки с одной или несколькими ковалентно связанными длинноцепочечными ацильными группами типичны для внутреннего монослоя. Кавеолин особенно часто встречается в вогнутых внутрь рафтах, называемых кавеолами (рис. 11-21). Белки с присоединенными пренильными группами (такие как Ras; см. доп. 12-2) имеют тенденцию к выталкиванию из рафтов. б) В данной искусственной мембране (созданной на поверхности слюды) из холестерина, синтетического фосфолипида диолеилфосфатидилхолина и GPI-связанного белка плацентарной щелочной фосфатазы наибольшая толщина наблюдается в области рафтов (поданным атомно-силовой микроскопии; см. доп. 11-1). Рафты выдаются из «океана» двойного липидного слоя (темная поверхность в верхней части верхнего монослоя); резкие пики соответствуют GPI-связанным белкам. Обратите внимание, что эти пики обнаруживаются почти исключительно в области рафтов.
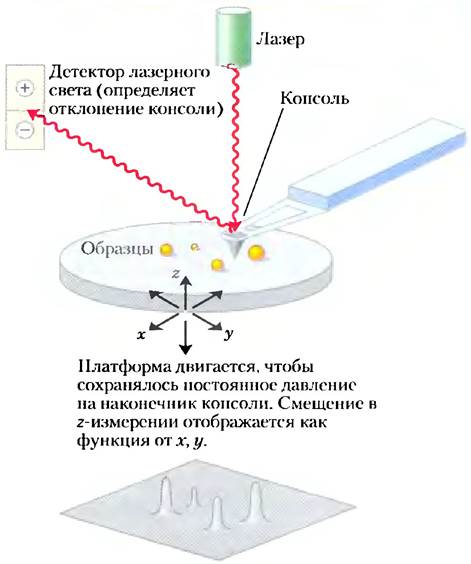
Дополнение 11-1. МЕТОДЫ. Атомно-силовая микроскопия для визуализации мембранных белков
В атомносиловой микроскопе (АСМ) на подвижной консоли закреплен микроскопический зонд с острым наконечником (рис. 1). Электростатические и ван-дер- ваальсовы взаимодействия между наконечником и образцом обусловливают силу, которая передвигает зонд вверх и вниз (в z-измерении) но мере того, как он проходит по образцу, отслеживая его рельеф. С помощью лазерного луча, отраженного от консоли, фиксируется движение даже па столь малое расстояние, как 1 А. В одном из типов атомно-силового микроскопа действующая па зонд сила поддерживается постоянной (порядка никоньютонов) с помощью контура с обратной связью, который заставляет платформу с образцом подниматься или опускаться, чтобы сила оставалась неизменной. Серия сканирований в направлениях x и у (в плоскости мембраны) дает трехмерную контурную карту поверхности образца с разрешением, близким к размеру атомов: 0,1 нм в вертикальном измерении; 0,5-1.0 им в латеральных направлениях. С помощью этой методики были визуализированы рафты, показанные на рис. 11-20, б.
Рис. 1
В благоприятных случаях АСМ можно использовать для изучения отдельных молекул мембранных белков. Одиночные молекулы бактериородоп- сина в пурпурных мембранах бактерии Halobacterium salinarium (рис. 11-9) видны как высокоупорядоченные структуры (рис. 2, а). При совмещении с помощью компьютера нескольких изображений отдельных единиц реальные части изображения усиливают друг друга, а шум исключается путем усреднения; на выходе получается изображение белка с высоким разрешением (вставка на рис. 2, а). При наблюдении как будто бы извне клетки реконструированный в липидных бислоях очищенный акванорип Е. соli демонстрирует в АСМ отчетливые детали пери плазматических доменов белка (рис. 2, б). Кроме того, АСМ показывает, что комплекс F0 (движущий протоны механизм АТР-синтазы хлоропластов; рис. 19-64) составлен из многих субъединиц, расположенных в виде круга (на рис. 2, в показано 14 субъединиц).
Рис. 2

Липидные рафты в высшей степени обогащены интегральными мембранными белками двух классов; одни заякорены в мембране двумя ковалентно связанными длинноцепочечными насыщенными жирными кислотами (две пальмитоильные группы или пальмитоильная и миристоильная группы), другие посредством GPI (рис. 11-14). Вероятно, эти липидные «якори», такие как ацильные цепи сфинголипидов, образуют более стабильные связи с холестерином и длинпоцепочечными ацильными группами в рафтах, чем с окружающими фосфолипидами. (Можно отметить, что другие связанные с липидами белки с ковалентно присоединенными изопренильными группами (например, фарсенильными) не обнаруживают предпочений в заякоривапии па внешнем монослое сфинголипид/холсстсриновых рафтов (рис. 11-20, а).) Домены «плота» и «моря» в плазматической мембране разделены нежестко; мембранные белки могут двигаться в липидные рафты внутрь и из рафтов наружу за время порядка нескольких секунд. Но в течение времени порядка нескольких микросекунд — масштаб, близкий тому, за который происходят многие опосредованные мембраной биохимические процессы, многие из этих белков могут считаться содержимым рафта.
Мы можем оценить долю поверхности клетки, занятой рафтами, по фракции плазматических мембран, которая устойчива к солюбилизации детергентами. В некоторых случаях она может составлять 50%: «плоты» покрывают половину «океана» (рис. 11-20, б). Косвенные измерения в культуре фибробластов позволяют грубо оцепить диаметр отдельного рафта в 50 нм, что соответствует путчу, содержащему несколько тысяч сфинголипидов и, возможно, от 10 до 50 мембранных белков. Поскольку большинство клеток экспрессируют более 50 разных видов белков плазматической мембраны, вероятно, что отдельный рафт содержит только субпопуляцию мембранных белков и что эта сегрегация мембранных белков имеет функциональное значение. Для процесса, при котором взаимодействуют два мембранных белка, их присутствие в одном рафте очень сильно увеличивает вероятность столкновения. Некоторые мембранные рецепторы и сигнальные белки, по-видимому, обособляются вместе в мембранных рафтах. Эксперименты показывают, что передачу сигнала через эти белки можно прервать с помощью манипуляций, которые выводят холестерин из плазматической мембраны и разрушают липидные рафты.
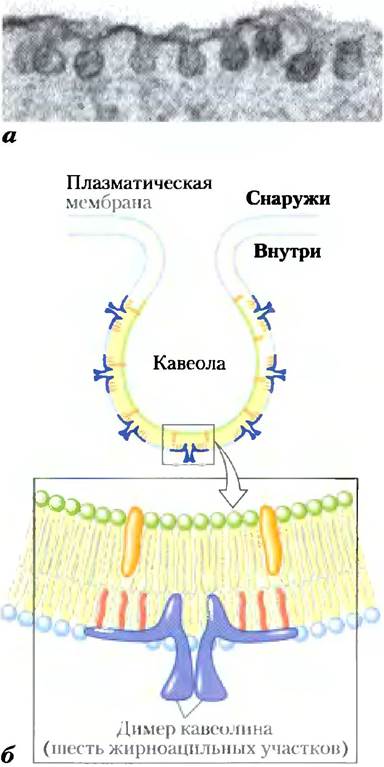
Кавеолин — это интегральный мембранный белок, содержащий два глобулярных домена, соединенных через петлеобразный гидрофобный домен, связывающий белок с цитоплазматическим монослоем плазматической мембраны. Три пальмитоильных группы, присоединенных к карбоксильному концевому глобулярному домену, обеспечивают его дополнительное заякоривание на мембране. Кавеолин (на самом деле это семейство родственных кавеолинов) связывает холестерин с мембраной, и присутствие кавеолина заставляет связанный с ним липидный бислой изгибаться внутрь, образуя кавеолы («маленькие полости») на поверхности клетки (рис. 11-21). Кавеолы — необычные рафты: в них есть оба монослоя — цитоплазматический монослой, из которого выступает глобулярный домен кавеолина, и внешний монослой — типичный сфинголипид-холсстериновый рафт с заякоренными через GPI белками. Кавеолы участвуют в разнообразных клеточных функциях, включая мембранный перенос внутри клеток и преобразование внешних сигналов в клеточные ответы. Рецепторы инсулина и других факторов роста и некоторые GTP-связывающие белки и про- теинкиназы, участвующие в передаче трансмембранных сигналов, по-видимому, локализованы в рафтах и, возможно, в кавеолах. В гл. 12 мы обсудим возможную роль рафтов в передаче сигналов.
Рис. 11-21. Кавеолин обусловливает вогнутые изгибы мембран. Кавеолы - это маленькие впячивания плазматической мембраны, как видно на электронной микрофотографии поверхности адипоцита, снятого с электронноплотным маркером (а), б) Каждый мономер кавеолина содержит центральный гидрофобный домен и три длинноцепочечные ацильные группы (показаны красным), которые удерживают молекулу на внутренней стороне плазматической мембраны. Когда несколько димеров кавеолина концентрируются на мембране в пределах маленькой области (рафте), они заставляют липидный бислой изогнуться, образуя кавеолу. Молекулы холестерина в бислое показаны оранжевым.
Изгибание и слияние мембран являются ключевым событием для многих биологических процессов
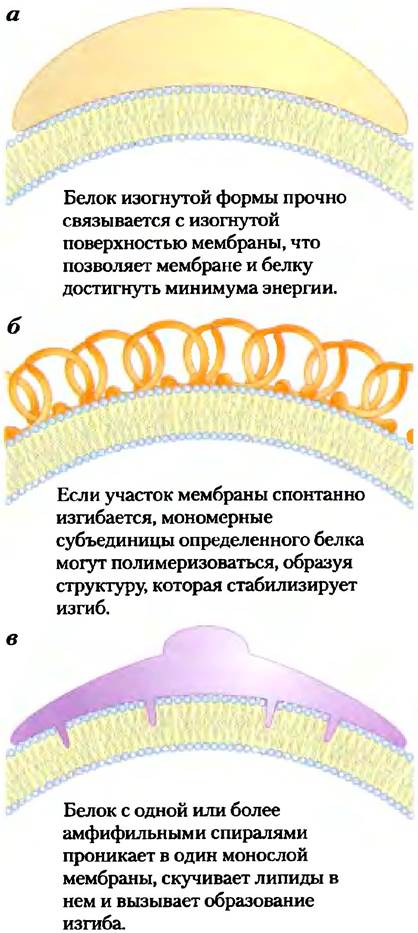
Не только кавеолин способен вызывать изгибы мембраны. Способность изгибаться лежит в основе одного из наиболее замечательных свойств биологических мембран — возможности слияния с другой мембраной без потери целостности. Хотя мембраны стабильны, их ни в коем случае нельзя считать статичными. Внутри эндомембранной системы эукариот (которая включает ядерную мембрану, эндоплазматический ретикулум, аппарат Гольджи и разнообразные малые везикулы) мембранные компартменты непрерывно реорганизуются. Везикулы отпочковываются от эндоплазматического ретикулума для того, чтобы переносить вновь синтезированные липиды и белки в другие органеллы и плазматическую мембрану. Экзоцитоз, эндоцитоз, клеточное деление, слияние сперматозоида и яйцеклетки, проникновение окруженного мембраной вируса в клетку хозяина — все эти процессы включают перестройку мембран, при которой происходит слияние двух мембранных сегментов без потери целостности (рис. 11-22). Большинство таких процессов начинается с небольшого искривления мембраны. На рис. 11-23 изображены три механизма, в соответствии с которыми может происходить искривление мембраны. Белок изогнутой формы может связываться с двойным липидным слоем и вызвать его искривление; движущей силой, увеличивающей изгиб мембраны, является энергия связывания. В другом варианте несколько субъединиц каркасного белка объединяются в изогнутый надмолекулярный комплекс и стабилизируют изгибы, спонтанно образующиеся в двойном слое. Кроме того, одна или несколько гидрофобных спиралей определенного белка могут проникать сквозь одну из поверхностей двойного слоя, расширяя ее относительно поверхности другой стороны двойного слоя, способствуя образованию изгиба.
Рис. 11-22. Слияние мембран. Слияние двух мембран — центральное событие в разнообразных процессах, в которых участвуют как мембраны органелл, так и плазматические мембраны.
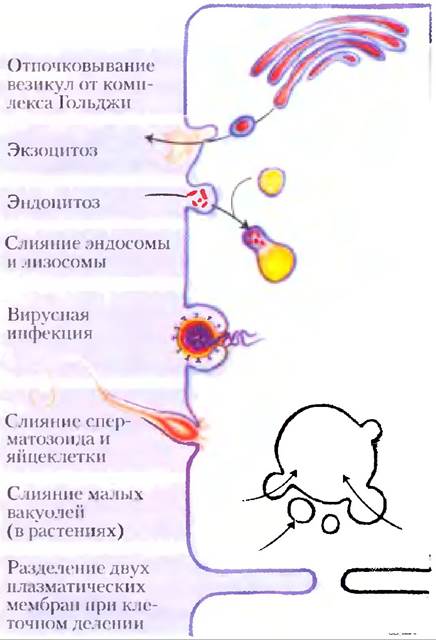
Рис. 11-23. Три модели изгибания мембраны, вызванного белками.
Специфическое слияние двух мембран требует, чтобы (1) они распознали друг друга; (2) их поверхности близко сошлись, для чего необходимо удаление молекул воды, обычно связанной с полярными «головками» липидов; (3) чтобы их бислойная структура локально разрушилась, обеспечивая слияние наружных монослоев каждой мембраны (полуслияние); и (4) чтобы их бислои слились, образуя один непрерывный бислой. Эндоцитоз с участием рецепторов или регулируемая секреция требует также, чтобы (5) процесс слияния запускался в надлежащее время или в ответ на специфический сигнал. Интегральные белки, называемые белками слияния, опосредуют эти события, осуществляя специфическое распознавание и кратковременную локальную деформацию бислойной структуры, что способствует слиянию мембран (заметим, что эти белки слияния не имеют отношения к продуктам двух слившихся генов, тоже называемым белками слияния, о которых говорилось в гл. 9).
Рис. 11-24. Слияние мембран в процессе высвобождения нейромедиатора в синапсе. Мембрана секреторной везикулы (секреторного пузырька) содержит v-SNAREсинаптобревин (красный цвет). Мембрана-мишень (плазматическая) содержит t-SNARE синтаксин (голубой цвет) и SNAР25 (фиолетовый). Когда локальное увеличение [Са2+] подает сигнал к высвобождению нейромедиатора, v-SNARЕ, SNАР25 и t-SNARE взаимодействуют, образуя переплетенный пучок из четырех α-спиралей. Это стягивает две мембраны вместе и локально разрушает бислой, что приводит к слиянию мембран и высвобождению нейромедиатора.
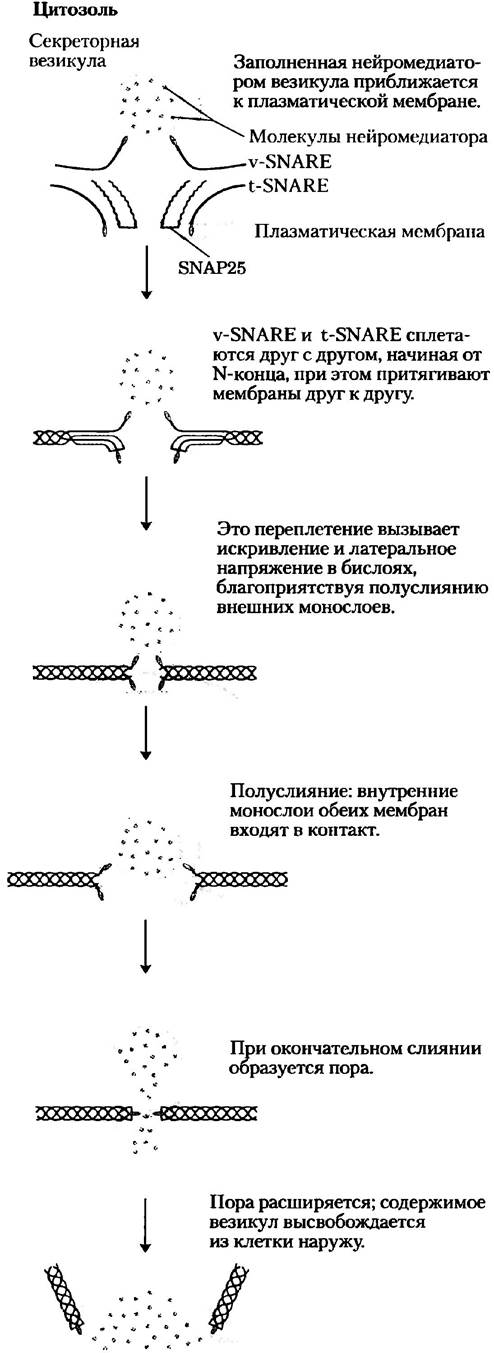
Особенно хороню изучено слияние мембран, происходящее в синапсах, где внутриклеточные везикулы, нагруженные нейромедиаторами, сливаются с плазматической мембраной. В этом процессе участвует семейство белков, называемых SNARE (рис. 11-24). Белки SNARE цитоплазматической стороны внутриклеточных везикул называются v-SNARE; белки в мембранах- мишенях, с которыми везикулы сливаются (плазматические мембраны в случае экзоцитоза), — это t-SNARE. Кроме того, участниками событий являются два других белка, SNAP25 и SNF. В процессе слияния мембран v-SNARE и t-SNARE связываются друг с другом и претерпевают структурную перестройку, в результате которой возникают пучки длинных тонких нитей, образованных из спиралей v-SNARE и t-SNARE и двух спиралей SNAP25 (рис. 11-24). Два белка SNARE сначала взаимодействуют своими концами, затем соединяются (как застежка-«молния»), образуя пучок спиралей. Это структурное изменение притягивает две мембраны для вступления в контакт и инициирует слияние их липидных бислоев.
Комплекс SNARE и SNAP25 является мишенью действия сильного токсина Clostridium botulinum — протеазы, которая расщепляет специфические связи в этих белках, препятствуя передаче нервных импульсов и вызывая гибель организма. Из-за очень высокой специфичности к этим белкам очищенный токсин ботулизма используется в качестве мощного инструмента для детального постадийного анализа механизма высвобождения нейромедиатора in vivo и in vitro.
Некоторые интегральные белки — посредники в межклеточных взаимодействиях и адгезии
Несколько семейств интегральных белков в плазматической мембране образуют специфические места связывания между клетками или между клеткой и белками внеклеточного матрикса. Интегрины — это белки поверхностной адгезии, которые опосредуют реакцию клетки на содержимое внеклеточного матрикса и другие клетки, не исключая и некоторые патогены. Кроме того, интегрины проводят сигнал через плазматическую мембрану в обе стороны, объединяя таким образом информацию снаружи и внутри клетки. Все интегрины представляют собой гетеродимерные белки (в них две разные субъединицы, α и β), заякоренные на плазматической мембране с помощью единичной трансмембранной спирали. Большие внеклеточные домены α- и β-субъединиц соединяются, образуя специфический связывающий участок для внеклеточных белков (таких как коллаген и фибронектин), который содержит одно общее звено, определяющее связывание интегринов, — последовательность Arg-Glу-Аsр (RGD). Более подробно сигнальные функции интегринов мы будем обсуждать в гл. 12 (с. 641).
Следующий тип мембранных белков, участвующих в поверхностной адгезии, — кадгерины; они взаимодействуют с идентичными кадгеринами в прилегающих клетках. Иммуноглобулин-подобные белки могут участвовать как во взаимодействиях с идентичными им партнерами в другой клетке, так и с интегрином соседних клеток. Селектины содержат внеклеточные домены, которые в присутствии Са2+ связывают специфические полисахариды, находящиеся на поверхности прилежащей клетки. Селектины присутствуют прежде всего в различных типах клеток крови и в клетках эндотелия кровеносных сосудов (рис. 7-33). Они играют существенную роль в процессах свертывания крови.
Интегральные белки участвуют во многих других клеточных процессах. Они служат транспортерами и ионными каналами (это обсуждается в разд. 11.3), функционируют как рецепторы гормонов, нейромедиаторов и факторов роста (гл. 12). Они играют центральную роль в окислительном фосфорилировании и фотосинтезе (гл. 19), в распознавании клеток в иммунной системе (клетка- клетка и антиген-клетка; гл. 5). Интегральные белки также активно участвуют в слиянии мем
бран, которое сопровождает экзоцитоз, эндоцитоз и вход многих вирусов в клетки хозяина.
Краткое содержание раздела 11.2 Динамика мембран
■ Липиды в биологической мембране могут существовать в жидко-упорядоченном или жидко-разупорядоченном состоянии; в последнем случае тепловое движение ацильных цепей делает внутренность бислоя жидкой. На текучесть влияют температура, состав жирных кислот и содержание стеринов.
■ Диффузия липидов по типу «флип-флоп» между внешним и внутренним монослоями мембраны — очень медленный процесс, за исключением тех случаев, когда его катализируют специфические флиппазы, флоппазы или скрамблазы.
■ Липиды и бачки могут двигаться латерально в плоскости мембраны, но такая подвижность ограничивается взаимодействиями мембранных белков с внутренними структурами цитоскелета и взаимодействиями липидов с липидными рафтами. К одному из классов липидных рафтов относятся рафты, которые состоят из сфинголипидов и холестерина с включением мембранных белков, связанных посредством GPI или длинноцепочечных ацильных групп.
■ Кавеолин — интегральный мембранный белок, который связывается с внутренним монослоем плазматической мембраны, заставляя ее искривляться с образованием кавеолы, вероятно, участвующей в мембранном транспорте и передаче сигналов.
■ Интегрины — трансмембранные белки плазматической мембраны, которые служат как для связывания клеток друг с другом, так и для передачи сигналов между внеклеточным матриксом и цитоплазмой.
■ Специфические белки опосредуют слияние двух мембран, которое сопровождает такие процессы, как внедрение вирусов, а также эндоцитоз и экзоцитоз.