ОСНОВЫ БИОХИМИИ ЛЕНИНДЖЕРА - ТОМ 1. ОСНОВЫ БИОХИМИИ СТРОЕНИЕ И КАТАЛИЗ - 2011
ЧАСТЬ I. СТРОЕНИЕ И КАТАЛИЗ
12. БИОСИГНАЛИЗАЦИЯ
12.9. Сигнализация у микроорганизмов и растений
Многое из того, что мы рассказали здесь о сигнализации, касается тканей млекопитающих или культур клеток, полученных из таких тканей. Бактерии, археи, эукариотические микроорганизмы и сосудистые растения тоже должны отвечать на множество внешних сигналов, таких как кислород, питательные вещества, свет, вредные химические вещества и т. д. Теперь перейдем к краткому рассмотрению видов сигнальных систем, которые используют микроорганизмы и растения.
Сигнализация у бактерий включает фосфорилирование в двухкомпонентной системе
Бактерия Escherichia coli реагирует на многие питательные вещества в окружающей среде, в том числе на сахара и аминокислоты, перемещением по направлению к ним, которое осуществляется с помощью одного или нескольких жгутиков. Мембранные белки содержат связывающие домены на внешней стороне плазматической мембраны, с ними связываются специфические аттрактанты (сахара или аминокислоты; рис. 12-30). Связывание лиганда приводит к тому, что другой домен на внутренней стороне плазматической мембраны фосфорилирует сам себя по остатку His. Затем этот первый компонент двухкомпонентной системы — рецепторная гистидинкиназа —катализирует перенос фосфорильной группы с остатка His на остаток Asp во втором, растворимом белке — регуляторе ответа; этот фосфопротеин перемещается к основанию жгутика, перенося сигнал от мембранного рецептора. Жгутик приводится в движение роторным мотором, который может перемещать клетку в окружающей среде или заставляет ее останавливаться в зависимости от направления вращения мотора. Полученная от рецептора информация позволяет клетке определить, движется она по направлению к источнику аттрактанта или от него. Если движение происходит по направлению к аттрактанту, регулятор ответа командует клетке продолжать движение по прямой; если движение происходит от источника аттрактанта, клетка мгновенно поворачивает, получая новое направление. Повторение такого поведения приводит к движению по случайной траектории в направлении увеличивающейся концентрации аттрактанта.
Рис. 12-30. Двухкомпонентный механизм сигнализации при бактериальном хемотаксисе. Когда лиганд-аттрактант А связывается с рецепторным доменом мебранносвязанного рецептора, протеингистидинкиназа в цитозольном домене (компонент 1) активируется и аутофосфорилируется по остатку His. Эта фосфорильная группа переносится затем на остаток Asp компонента 2 (или отдельный белок, или другой домен рецепторного белка). После фосфорилирования по Asp компонент 2 передвигается к основанию жгутика, где он определяет направление вращения мотора жгутика.
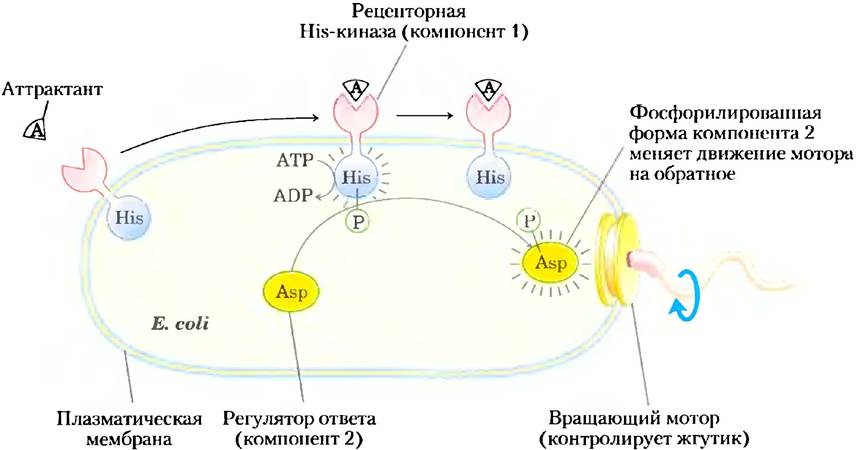
Используя эту основную двухкомпонентную систему, Е. coli реагирует не только на сахара и аминокислоты, но также на кислород, перепады температуры и другие факторы окружающей среды. Двухкомпонентная система обнаружена у многих бактерий, как у грамположительных, так и у грамотрицательных, а также у архей, простейших и грибов. Очевидно, что этот механизм сигнализации развился на ранней стадии в ходе клеточной эволюции и впоследствии был сохранен.
Различные сигнальные системы, используемые животными клетками, имеют аналоги также у прокариот. По мере того как становились известны полные геномные последовательности все большего числа различных бактерий, исследователи открывали гены, которые кодируют белки, аналогичные серин- или тирозинкиназам, Ras-подобные белки, регулируемые связыванием с GТР, и белки с SH3-доменами. Рецепторные Туг-киназы не обнаружены у бактерий, но в некоторых бактериальных белках встречаются остатки Ⓡ-Туr, поэтому должен существовать и фермент, который фосфорилирует остатки Туr.
Сигнальные системы растений содержат компоненты, которые используются микроорганизмами и млекопитающими
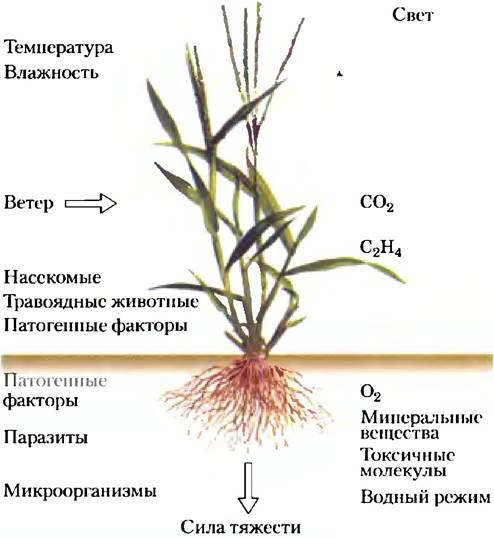
Подобно животным, сосудистые растения должны иметь средства коммуникации между тканями для того, чтобы координировать и регулировать рост и развитие; чтобы адаптироваться к кислородному режиму, питательным веществам, свету и температуре; чтобы предупреждать о присутствии вредных химических веществ и повреждающих патогенных факторов (рис. 12-31). С тех пор, когда разошлись растительные и животные ветви эукариот, миновал по крайней мере миллиард лет эволюции, это отразилось на механизмах сигнализации: некоторые механизмы сигнализации у растений консервативны, т. с. подобны механизмам, присущим животным (протеинкиназы, адаптерные белки, циклические нуклеотиды, электрогенные ионные насосы и регулируемые ионные каналы); некоторые сходны с бактериальными двухкомпонентными системами; другие свойственны исключительно растениям (например, механизмы, воспринимающие свет) (табл. 12-7). Геном всесторонне изученного растения Arаbidopsis thaliana, например, кодирует около 1000 белковых Ser/Thr-киназ, в том числе около 60 МАРК и около 400 ассоциированных с мембраной рецепторных киназ, которые фосфорилируют остатки Ser или Thr; множество протеинфосфатаз; адаптерные белки, собирающие другие белки в сигнальные комплексы; ферменты для синтеза и деградации циклических нуклеотидов; и 100 или более ионных каналов, включая примерно 20 ргулируемых циклическими нуклеотидами. Присутствуют также инозитфосфолипиды и киназы, которые превращают их один в другой путем фосфорилирования инозитольных полярных групп.
Таблица 12-7. Сигнальные компоненты млекопитающих, растений или бактерий
Сигнальный компонент |
Млекопитающие |
Растения |
Бактерии |
Ионные канаты |
+ |
+ |
+ |
Электрогенные ионные насосы |
+ |
+ |
+ |
Двухкомпонентпые His-киназы |
+ |
+ |
+ |
Аденилатциклаза |
+ |
+ |
+ |
Гуанилатциклаза |
+ |
+ |
? |
Рецепторные протеинкиназы (Ser/Thr) |
+ |
+ |
? |
Са2+ как вторичный мессенджер |
+ |
+ |
? |
Са2+-капалы |
+ |
+ |
? |
Кальмодулин, СаМ-связывающий белок |
+ |
+ |
- |
Каскад МАРК |
+ |
+ |
- |
Канаты, управляемые циклическим нуклеотидом |
+ |
+ |
- |
IP3-регулирусмые Са2+-капаты |
+ |
+ |
- |
Фосфатидилнозиткиназы |
+ |
+ |
- |
GPCR |
+ |
+/- |
+ |
Тримерные G-белки |
+ |
+/- |
- |
PI-специфическая фосфолипаза С |
+ |
? |
- |
Тирозинкиназные peцепторы |
+ |
? |
- |
SН2-домены |
+ |
? |
? |
Ядерные рецепторы стероидов |
+ |
- |
- |
Протеинкиназа А |
+ |
- |
- |
Протеинкиназа G |
+ |
- |
- |
Рис. 12-31. Некоторые стимулы, вызывающие ответы в растениях.
Однако некоторые виды сигнальных белков, широко распространенные в тканях животных, отсутствуют у растений или представлены только небольшим числом генов, например, по-видимому, отсутствуют циклонуклеотидзависимые протеинкиназы (РКА и РКG). Гены гетеротримерных G-белков и протеинтирозинкиназы в гораздо меньшей степени заметны в геноме растений, а гены GРCК (самое большое семейство в геноме человека, -1000 генов) очень бедно представлены в растительном геноме. Ядерные рецепторы стероидов, связанные с ДНК, не обнаружены и, возможно, вообще отсутствуют в растениях. Хотя в растениях отсутствует большинство широко распространенных общих светочувствительных механизмов, которые имеются у животных (родопсин с ретиналем в качестве пигмента), там есть широкий набор других воспринимающих свет механизмов, не обнаруженных в тканях животных, например, фитохромы и криптохромы (гл. 19).
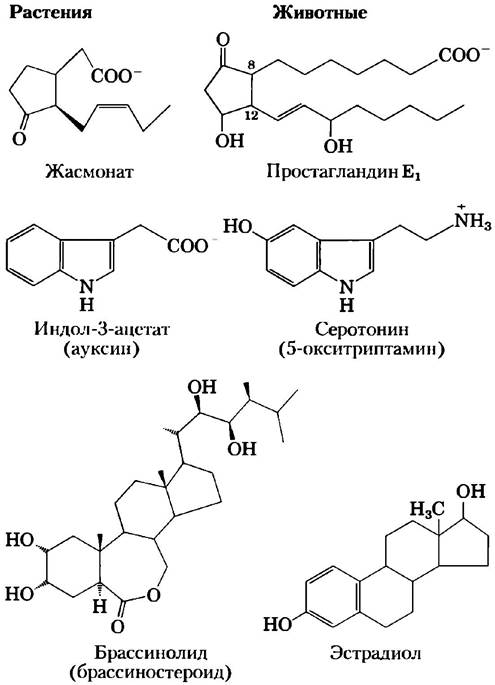
Классы соединений, продуцирующие сигналы в растениях, сходны с некоторыми сигнальными молекулами у млекопитающих (рис. 12-32). Вместо простагландинов растения образуют жасмонат; вместо стероидных гормонов — брассиностероиды. Сигнальными веществами в растениях служат около 100 разнообразных низкомолекулярных пептидов. И растения, и животные используют в качестве сигнальных производные ароматических аминокислот.
Рис. 12-32. Структурное сходство между растительными и животными сигнальными веществами. Растительные сигнальные вещества жасмонат, индол-3-ацетат и брассинолид похожи на сигнальные вещества млекопитающих простагландин Е1, серотонин и эстрадиол.
Растения обнаруживают этилен с помощью двухкомпонентной системы и каскада МАРК
Рецепторы газообразного растительного гормона этилена (СН2=СН2) по первичной структуре сходны с рецепторными His-киназами бактериальных двухкомпонентных систем и, вероятно, развились из них в процессе эволюции. У р. Аrаbidopsis двухкомпонентная сигнальная система состоит из единственного интегрального белка эндоплазматического ретикулума (а не плазматической мембраны). Этилен диффундирует в клетку через плазматическую мембрану и проникает в ЭР. Первый компонент в нисходящем ряду, затронутый этиленовой сигнализацией, — это протеин-Ser/Тhr-киназа (СТR-1; рис. 12-33) с последовательностью, гомологичной Raf — протеинкиназе, которая у млекопитающих начинает каскад МАРК в ответ на действие инсулина (ср. рис. 12-15). У растений в отсутствие этилена киназа СТR активна и ингибирует каскад МАРК, препятствуя транскрипции этилен-чувствительных генов. Действие этилена инактивирует киназу СТR-1, активируя тем самым каскад МАРК, что приводит к активации фактора транскрипции ЕINЗ. Активный EIN3 стимулирует синтез второго фактора транскрипции (ERF1), который в свою очередь активирует транскрипцию ряда этилен-чувствительных генов; продукты этих генов влияют на процессы, начиная с развития семян и кончая созреванием плодов.
Рис. 12-33. Механизм передачи сигнала при обнаружении этилена растениями. Этиленовый рецептор в плазматической мембране (розовый цвет) представляет собой двухкомпонентную систему из одного белка, который имеет рецепторный домен (компонент 1) и домен- регулятор ответа (компонент 2). Рецептор регулирует (каким образом — мы пока не понимаем) активность CTR1 — протеинкиназы, подобной МАРКК киназам и, вероятно, являющейся, частью каскада МАРК. СТR1 — отрицательный регулятор ответа на этилен; когда СTR1 неактивна, этиленовый сигнал проходит через продукт гена EIN2 (полагают, что это белок ядерной оболочки), который каким-то образом вызывает увеличение синтеза ERF1 — фактора транскрипции; ЕRF1 в свою очередь стимулирует экспрессию белков, специфических для ответа на действие этилена.
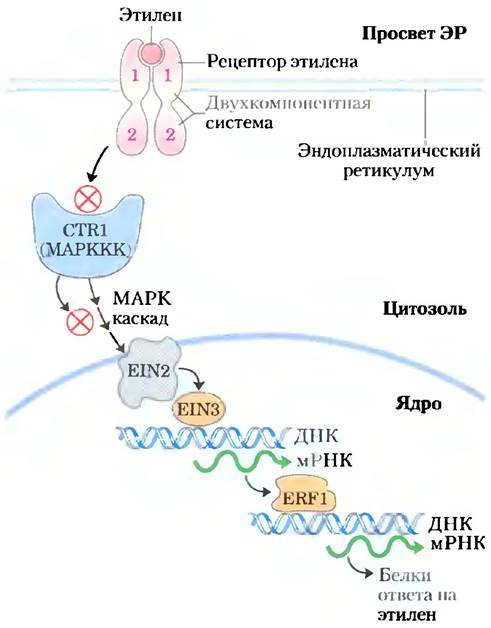
Очевидно, возникшая из бактериальной двухкомпонентной сигнальной системы этиленовая система у р. Arabidopsis отличается тем, что His-киназная активность, которая у бактерий является компонентом 1, несущественна при передаче сигнала у р. Arabidopsis.
Рецептороподобные протеинкиназы осуществляют передачу сигналов от пептидов и брассиностероидов
Одним общим мотивом при передаче сигналов в растениях является участие рецептороподобных протеинкиназ (RLK от англ. receptorlike kinases), содержащих один спиральный сегмент в плазматической мембране, который осуществляет связь рецепторного домена на внешней стороне мембраны с иротеин-Scr/Thr- киназой на цитоплазматической стороне. Этот тип рецептора участвует в механизме защиты при инфицировании бактериальным патогеном (рис. 12-34, а). Сигналом для генов, необходимых для защиты против инфекции, служит пептид flg22, высвобождаемый при распаде флагеллина — основного белка бактериального жгутика. Связывание flg22 с рецептором ELS2 у р. Arabidopsis вызывает димеризацию рецептора и его аутофосфорилирование по остаткам Ser и Thr, а последующим эффектом является активация каскада МАРК подобно тому, как это было описано выше для действия инсулина (рис. 12-15). Конечная киназа в этом каскаде активирует специфический фактор транскрипции, запускающий синтез белков защиты от бактериальной инфекции. Стадии между фосфорилированием рецептора и каскадом МАРК пока неизвестны. Для прекращения ответа на стимул с активным рецепторным белком связывается фосфопротеин-фосфатаза (КАРР), которая инактивирует его путем дефосфорилирования.
Рис. 12-34. Сходство между сигнальными путями, запускающими иммунные ответы у растений и животных, а) У растения Arabidopsis thaliana петид flg22, происходящий из жгутиков бактериального патогена, связывается с рецептором на плазматической мембране, вызывая образование из рецепторов димеров и запуская аутофосфорилирование цитозольного протеинкиназного домена по остаткам Ser и Thr (не Туr). Аутофосфорилирование активирует рецепторную протеинкиназу, которая затем фосфорилирует следующие в цепи белки. Активированный рецептор активирует также (неизвестно, каким образом) МАРККК. Итоговый киназный каскад приводит к фосфорилированию ядерного белка, который обычно ингибирует факторы транскрипции WRKY22 и 29, приводя к деградации ингибитора и давая возможность факторам транскрипции стимулировать экспрессию генов, имеющих отношение к иммунному ответу. б) У млекопитающих токсический бактериальный липополисахарид (ЛПС; см. рис. 7-30) обнаруживается рецепторами плазматической мембраны, которые связываются с растворимой протеинкиназой (IRAK) и активируют ее. Основной жгутиковый белок патогенной бактерии действует через похожий рецептор и активирует IRAK. Затем IRАК инициирует два разных каскада МАРК, которые заканчиваются в ядре, вызывая синтез белков, необходимых для иммунного ответа. Jun, Fos, NFkB — факторы транскрипции.
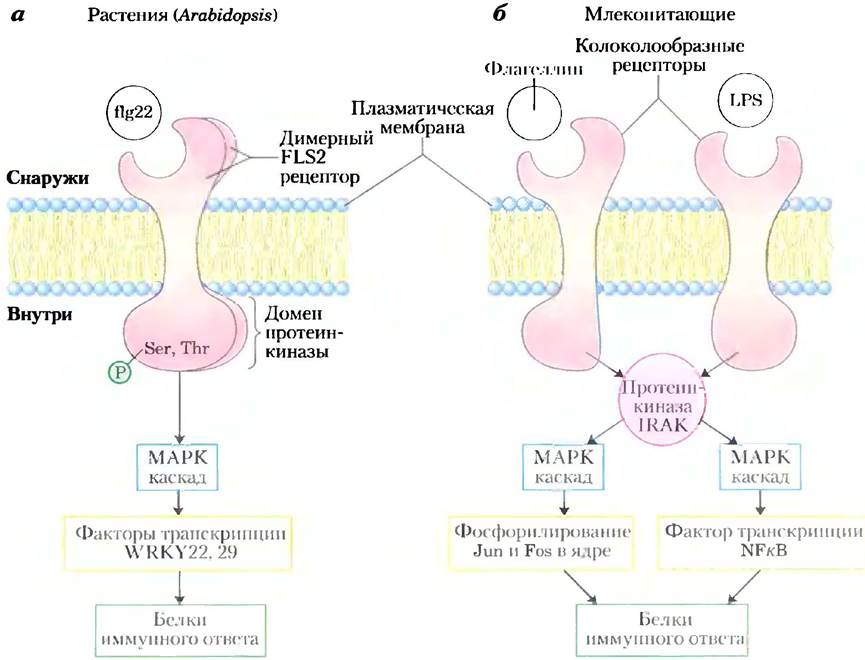
Каскад МАРК в защите растений против бактериальных патогенов поразительно похож на генетически детерминированный иммунный ответ у млекопитающих (рис. 12-34, б), запускаемый бактериальным липополисахаридом и опосредуемый Toll-подобными рецепторами (TLR, от мутанта р. Drosophila, который обычно называют «Toll» — от нем. сумасшедший; впоследствии TLR обнаружены у многих организмов и было показано их участие в эмбриогенезе). Другие мембранные рецепторы используют похожие механизмы активации каскада МАРК, активируя в конечном счете факторы транскрипции и вводя в действие гены, необходимые для защитной реакции.
Предполагают, что из нескольких сотен RLK в растениях большинство работают подобным образом: связывание лиганда индуцирует димеризацию и аутофосфорилирование, а активированные рецепторные киназы запускают последующие реакции путем фосфорилирования ключевых белков по остаткам Ser и Thr.
Краткое содержание раздела 12.9 Сигнализация у микроорганизмов и растений
■ Бактерии и эукариотические микроорганизмы обладают множеством разнообразных сенсорных систем, которые позволяют им «брать пробы» окружающей среды и реагировать на воздействия окружения. В двухкомпонентной системе рецепторная His-киназа воспринимает сигнал и аутофосфорилирует остаток His, затем фосфорилирует белок- регулятор ответа по остатку Asp.
■ Растения отвечают на многие стимулы окружающей среды и используют гормоны и факторы роста для того, чтобы координировать развитие и метаболическую активность своих тканей. Геном растений кодирует сотни сигнальных белков, в том числе и некоторые белки, очень похожие на используемые для передачи сигналов в клетках млекопитающих.
■ Двухкомпонентные механизмы сигнализации, широко распространенные у бактерий, обнаружены в измененной форме у растений, где служат для детекции химических сигналов и света.
■ Растительные рецептороподобные киназы (RLK) участвуют в восприятии множества стимулов, в том числе брассиностероидов, пептидов патогенных организмов и сигналов, участвующих в процессе развития. RLK аутофосфорилируют остатки Ser/Thr, затем активируют последующие белки, которые в некоторых случаях образуют каскад МАРК. Конечным результатом множества такого рода сигналов является усиление транскрипции специфических генов.