Основы биохимии Том 2 - А. Ленинджер 1985
Биоэнергетика и метаболизм
Метаболизм. Общий обзор
Краткое содержание главы
Организмы можно классифицировать исходя из их потребности в тех или иных источниках углерода. Автотрофам достаточно двуокиси углерода, а гетеротрофы должны получать углерод в виде каких-нибудь восстановленных органических соединений, таких, как глюкоза. Для многих автотрофных клеток, например для клеток зеленых растений, источником энергии служит солнечный свет; гетеротрофні получают необходимую им энергию в результате окисления органических пищевых веществ.
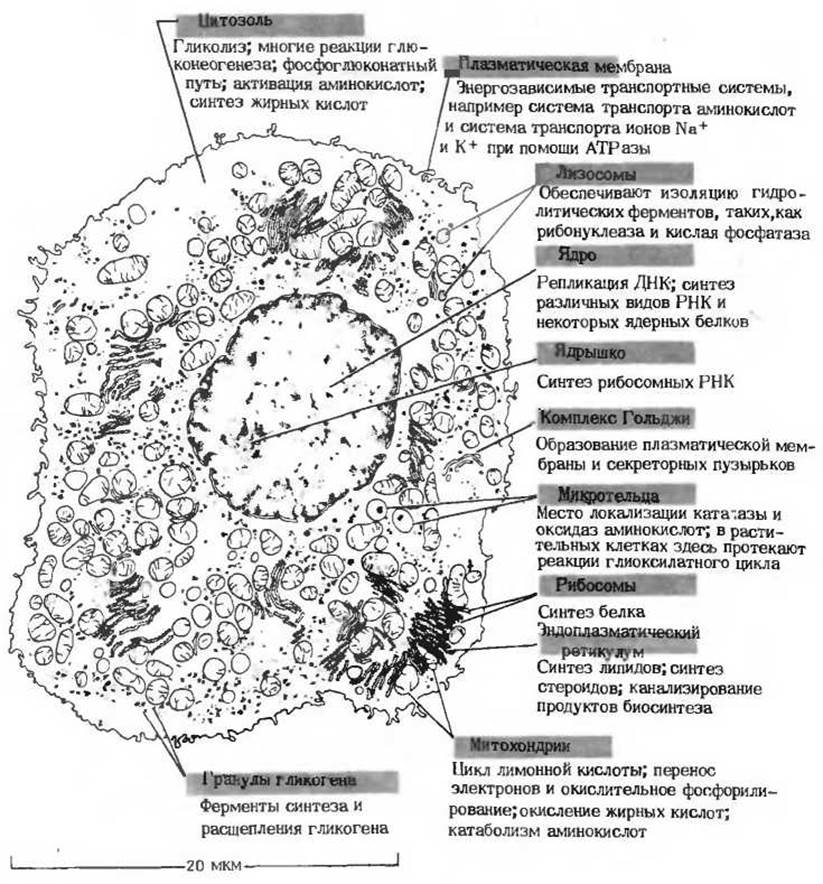
Рис. 13-23. Компартментализация некоторых важных ферментов и метаболических путей в клетке печени крысы. Электронная микрофотография, на основе которой выполнен этот рисунок, приведена в гл. 2 (см. рис. 2-7).
Метаболизм включает в себя катаболизм, или расщепление пищевых веществ, богатых энергией, и анаболизм, или биосинтез новых клеточных компонентов. В катаболических и анаболических процессах различают три главные стадии. На первой стадии катаболизма полисахариды, жиры и белки расщепляются под действием ферментов до своих строительных блоков, на второй стадии происходит окисление этих строительных блоков, в результате которого в качестве главного продукта образуется ацетил-СоА, а на третьей стадии ацетильная группа ацетил-СоА окисляется до СО2. Различные катаболические пути сливаются в один общий конечный путь, анаболические же пути расходятся, так что из небольшого числа предшественников образуется в конечном счете много различных продуктов. Соответствующие катаболические и анаболические пути неидентичны, т.е. отдельные их ферментативные этапы не совпадают; регулируются эти пути независимо; часто они локализованы в разных участках клетки. При распаде пищевых веществ часть высвобождающейся из них энергии запасается в форме аденозинтрифосфата (ATP). ATP служит переносчиком энергии от катаболических реакций к процессам, сопровождающимся потреблением энергии, таким, как биосинтез, сокращение или движение, перенос веществ через мембрану или передача генетической информации. Химическая энергия передается также от катаболических процессов к анаболическим в форме восстановительной способности - через восстановленный кофермент NADPH.
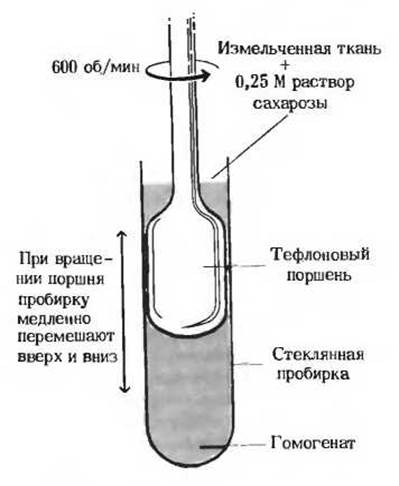
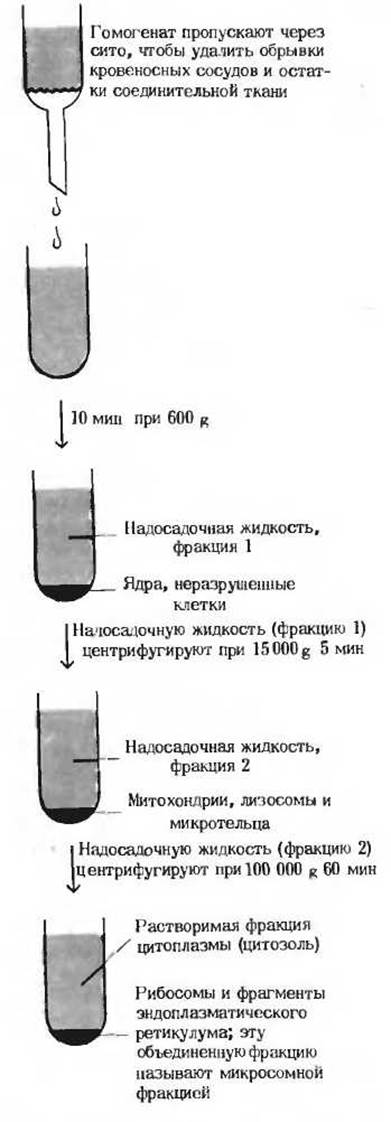
Рис. 13-24. Фракционирование клеточного экстракта методом дифференциального центри фугирования. Клеточная мембрана разрушается силами трения в гомогенизаторе с вращающимся поршнем. После удаления остатков соединительной ткани и обрывков кровеносных сосудов при помощи сита из нержавеющей стали клеточный экстракт центрифугируют несколько раз, постепенно увеличивая скорость вращения ротора.
Регуляция метаболизма осуществляется на трех уровнях: 1) при помощи аллостерических ферментов, 2) при помощи гормонов и 3) путем регулирования синтеза ферментов. Для анализа метаболических путей применяют экстракты клеток и тканей, из которых выделяют ферменты изучаемого метаболического пути и его промежуточные продукты. Широкие возможности для изучения метаболизма открывает использование мутантных микроорганизмов с генетическими дефектами, затрагивающими тот или иной метаболический путь (ауксотрофных мутантов). Весьма эффективен в этом отношении также метод изотопных меток. В эукариотических клетках ферменты различных метаболических путей разделены пространственно, т.е. находятся в разных органеллах - в ядрах, митохондриях или эндоплазматическом ретикулуме; из этих органелл их можно выделять для прямого изучения.
ЛИТЕРАТУРА
Книги
Colowick S. Р., Kaplan N. О. (eds.). Methods in Enzymology, Academic, New York, 1955. Многотомное издание, содержащее обзорные статьи по всем вопросам энзимологии и метаболизма, а также по методам исследований.
Dagley S.. Nicholson D. Е. Introduction to Metabolic Pathways, Wiley, New York, 1970. (Имеется перевод: Дэгли С., Николсон Д. Метаболические пути.-М.: Мир, 1973.) Сводка метаболических карт.
Roodyn D. В. (ed.). Subcellular Biochemistry, V. I—VII, Plenum, New York, 1972-1980. Многотомное издание, содержащее обзорные статьи по компартментализации различных видов биохимической активности в клетках.
Stonier R.Y.. Doudoroff М., Adelberg Е. A. The Microbial World, 4th ed, Prentice-Hall, Englewood Cliffs, N.J, 1976. (Имеется перевод: Стейниер P. и др. Мир микробов. В трех томах - М.: Мир, 1979.) Хороший учебник по общей микробиологии. Рассматриваются различные подгруппы автотрофных и гетеротрофных организмов; обсуждается существенный вклад микроорганизмов в общий обмен биомассы земного шара.
Статьи
Сентябрьский номер журнала «Scientific American» за 1970 г. (т. 223, № 3) целиком посвящен вопросам, касающимся биосферы. Особо можно рекомендовать следующие статьи, вошедшие в этот номер журнала:
Penman Н. L. The Water Cycle, р. 98.
Cloud Р., Gibor A. The Oxygen Cycle, p. 110. Bolin B. The Carbon Cycle, p. 124.
Delwiche C.C. The Nitrogen Cycle, p. 136. Brown L. R. Human Food Production as a Process in the Biosphere, p. 160.
Hall D. O. Photobiological Energy Conversion, FEBS Lett., 64, 6-16 (1976). Общая статья, рассматривающая фотосинтез с точки зрения потребностей человека в пище и энергии.
Woodwell G. М. The Carbon Dioxide Question, Sci. Am, 238, 34-43, January 1978. Обсуждаются возможные последствия повышения содержания СO2 в атмосфере.
Вопросы и задачи
1. Анализ метаболического пути. Анализ любого метаболического пути напоминает подведение баланса в приходно-расходной книге. В такую книгу заносят все произведенные в течение месяца операции (в соответствующую главу «Приход» или «Расход»), а в конце месяца определяют итог. Метаболический путь - это тоже ряд «операций», ряд последовательных химических превращений, из которых в конечном счете и слагается данный метаболический процесс. Подобно бухгалтерским операциям, химические превращения тоже можно охарактеризовать количественно и каждое из них может быть описано уравнением химического баланса. В интактных клетках такой «учет» - далеко не простое дело, поскольку промежуточные продукты данного метаболического пути вовлекаются иногда в другие метаболические процессы. Ниже приведены реакции, из которых складывается процесс ферментативного превращения глицеральдегид-3-фосфата в этанол (спиртовое брожение) в дрожжевых клетках. Каждое из приведенных здесь химических уравнений сбалансировано. Следует, однако, иметь в виду, что в лействительности реакции, из которых состоит данный метаболический путь, могут протекать и в иной последовательности.
Глицеральдегид-3-фосфат + Рі + NAD+→ 3-фосфоглицероилфосфат + NADH + Н + Фосфоенолпируват + ADP → Пируват + АТР
Этанол + NAD+→ Ацетальдегид + NADH + Н+
3-фосфоглицероилфосфат + ADP → 3-фосфоглицерат + АТР
2-фосфоглицерат ⇄ 3-фосфоглицерат
2-фосфоглицерат → Фосфоенолпируват + Н2О
Пируват → СО2 + Ацетальдегид
а) Используя все эти уравнения, записанные в словесной форме, и зная химическую структуру каждого промежуточного продукта, выведите и запишите последовательность химических превращений (метаболический путь), из которых состоит процесс расщепления глицеральдегид-3-фосфата до этанола.
б) Напишите суммарное уравнение этого процесса. Для этого вам придется поступить так, как если бы вы подбивали итог в приходно-расходной книге, т.е. суммировать все поступления и траты.
в) Изобразите данный метаболический путь так, как это сделано на рис. 13-4. Укажите, как связаны между собой отдельные части этого пути.
2. Циклический метаболический путь. Среди бактерии, принадлежащих к роду Pseudomonas, встречаются виды, способные использовать в качестве источника клеточного топлива оксалат (-ООС—СОО-) - соединение, крайне токсичное для большинства млекопитающих. Обмен оксалата у таких бактерий идет по циклическому пути, который схематически можно представить следующим образом:
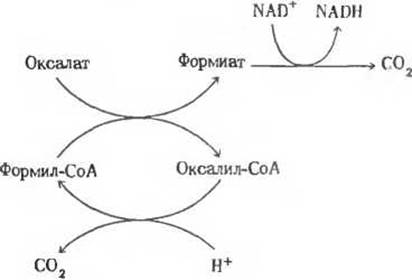
а) Изобразите данный метаболический путь в виде ряда уравнений химического баланса, в которых продукт одной реакции служит субстратом следующей. Напишите формулы соединений, участвующих в каждой из стадий.
б) Напишите сбалансированное суммарное уравнение процесса расщепления оксалата до СО2, пользуясь структурными формулами.
3. Включение 14С в аминокислоты. В 1955 г. Робертс и его сотрудники опубликовали результаты своих экспериментов по выращиванию Escherichia coli на среде, содержавшей в качестве единственного источника углерода равномерно меченную 14С-глюкозу. Исследователи рассчитывали таким способом пометить радиоактивным углеродом биомолекулы клетки. Оказалось, что при выращивании бактерий на 14С-глюкозе метка быстро включается во все аминокислоты. Если же в среду, содержащую 14С-глюкозу, добавляли немеченый гистидин (так что источников углерода было уже два: глюкоза и гистидин), то 14С включался во все аминокислоты, за исключением гистидина. Почему при отсутствии немеченого гистидина 14С включался в эту аминокислоту? Почему в среде, содержавшей немеченый гистидин, метка в гистидин не включалась? Что происходило: ингибирование по типу обратной связи или репрессия ферментов?
4. Регуляция обмена лактозы у Е. coli. Клетки Е. coli можно выращивать на простой среде, содержащей в качестве единственного источника углерода лактозу. Необходимым этапом обмена лактозы (а следовательно, и необходимым условием выживания бактерии) является гидролиз лактозы до моносахаридов глюкозы и галактозы, катализируемый ферментом ß-гaлактозидазой. Когда Е. coli выращивают на среде с лактозой, в каждой бактериальной клетке присутствует несколько тысяч молекул этого фермента (см. задачу 1), способных осуществлять гидролиз. Однако если единственным источником углерода в среде служит глюкоза или, например, глицерол, то в клетках обнаруживается не более 5-10 молекул ß-галактозидазы.
а) Каким образом регулируется обмен лактозы? Объясните.
б) Почему при замене в питательной среде лактозы на глицерол содержание ß-галактозидазы в клетках снижается? Почему оно не остается на прежнем уровне?
в) Если среда содержит в качестве единственного источника углерода метил-ß-галактозид, то клетки растут быстро и содержат тысячи молекул ß-галактозидазы. Если же единственным источником углерода является метил-а-галактозид, то клетки растут медленно и содержат совсем мало ß-галактозидазы. Объясните причины этих различий.
5. Сравнение катаболических и анаболических путей. Ниже на схеме изображено взаимопревращение глюкозы и фруктозо-1,6-дифосфата. В обмене углеводов эта последовательность реакций играет ключевую роль. Расщепление глюкозы представляет собой катаболический путь, а ее биосинтез из фруктозо-1,6-дифосфата- анаболический. Одни и те же гексозомонофосфаты служат промежуточными проектами того и другого пути. Однако, хотя пути эти очень схожи, между ними есть явные различия. Выявите их.
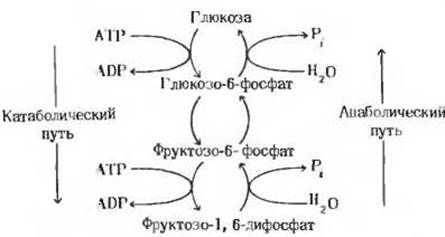
а) Напишите уравнения химического баланса для каждой из стадий катаболического пути. Напишите суммарное уравнение, представляющее собой результат сложения отдельных стадий.
б) Проделайте то же самое для анаболического пути.
в) Укажите различия между катаболическим и анаболическим путями, проявляющиеся в их суммарных уравнениях. Можно ли считать, что каждый из этих путей является простым обращением другого?
г) Чем обеспечивается направленность катаболизма глюкозы? Иными словами, что препятствует обращению этого процесса?
д) Возможно ли, чтобы один и тот же фермент катализировал и катаболическую, и анаболическую реакции взаимопревращения глюкозы и глюкозо-6- фосфата? Возможно ли это для взаимопревращения глюкозо-6-фосфата и фруктозо-6-фосфата?
6. Измерение радиоактивности. Количественное определение радиоактивных изотопов, обычно используемых в биологических исследованиях (3Н, 14С, 32Р и 35S), удобнее всего проводить при помощи жидкостного сцинтилляционного счетчика. За единицу радиоактивности (ее называют кюри, Ки; международное обозначение -Сі) принята активность, соответствующая 2,22-1012 распадам в 1 минуту (расп./мин). При использовании жидкостного сцинтилляционного счетчика обычно удается зарегистрировать в виде импульсов только часть общего числа распадов. Поэтому количество радиоактивности часто выражают не в виде числа распадов в минуту (расп./мин), а в виде числа реально регистрируемых данным счетчиком импульсов в минуту (имп/мин). Если известна эффективность счета, которая определяется по формуле то на основании числа регистрируемых импульсов в минуту можно определить число распадов в минуту.
![]()
Использование радиоактивной метки - очень ценный метод биохимических исследований, потому что по радиоактивности мы можем судить о концентрации данного химического соединения. Под удельной радиоактивностью соединения понимают активность на единицу массы, объема и т.д. (грамм, моль, эквивалент, миллилитр и т.д.). Пользуясь этим понятием, мы вовсе не предполагаем, что метку несет каждая молекула данного соединения. Единственное, что требуется, - это чтобы регистрируемое счетчиком число импульсов было пропорционально концентрации меченого соединения; удельная радиоактивность и эффективность счета служат при этом пересчетными коэффициентами.
а) От поставщика получен препарат 14С-глюкозы в 1 мл водного раствора. Общая радиоактивность препарата 250 мкКи, а его удельная радиоактивность 500 мКи/ммоль. Вычислите концентрацию глюкозы в полученной от поставщика 1-мл ампуле.
б) Сколько импульсов в минуту будет зарегистрировано. если из полученного раствора отобрать пробу (10 мкл), ввести ее в счетчик и измерить радиоактивность с эффективностью 70 %?
7. Измерение концентрации метионина внутри клетки. Когда клетки Е. coli выращивают на среде, в которой единственным источником серы служит 35SО2-4, все серусодержащие аминокислоты и белки содержат метку 35S. В одном из таких экспериментов Е. coli выращивали на среде, содержащей 0,85 мМ 35SO2-4. При измерении радиоактивности в 250-мкл пробе этой среды с 87%-ной эффективностью счета было зарегистрировано 4,50∙105 имп/мин. По достижении максимального роста культуры клетки отфильтровали и промыли холодной водой. Свободные (несвязанные) аминокислоты экстрагировали из клеток кипящей водой и разделили методом ионообменной хроматографии. Экстракт из 1,85 г влажных клеток при измерении с 82%-ной эффективностью имел радиоактивность 3,2∙105 имп/мин L-35S-метионина. Вычислите концентрацию свободного L-метионина внутри клетки, исходя из предположения, что во влажных клетках 80% составляет вода и 20%-сухой остаток.