Биохимия - Химические реакции в живой клетке Том 1 - Д. Мецлер 1980
Как молекулы соединяются друг с другом
Упаковка макромолекул
Спиральные структуры
Если угол, образуемый двумя субъединицами при связывании а с j, несколько отличается от угла, соответствующего замкнутому кольцу, то образуется структура типа спирали, изображенная на рис. 4-6, Б. На один виток спирали может приходиться как целое число субъединиц, так и не целое (как в спиральной структуре, показанной на рисунке). Каждая последующая субъединица присоединяется к предыдущей за счет тех же гетерологических контактов типа aj, однако в этом случае могут иметь место и другие взаимодействия. Если поверхности субъединиц комплементарны и их геометрия благоприятствует образованию дополнительных контактов, то группы, расположенные в разных частях молекулы (например, b и k), могут соединиться друг с другом, образуя другую гетерологическую связь. Возможно образование и третьей гетерологической связи, cl, между двумя другими участками поверхностей субъединиц. Если контакты aj, bk и cl достаточно прочны (т. е. если площади комплементарных поверхностей субъединиц достаточно велики, а сами поверхности высококомплементарны), то могут образовываться чрезвычайно прочные структуры из микротрубочек (например, жгутики у эукариот, рис. 1-5). Когда взаимодействия не столь сильны, то образуются часто обнаруживаемые в клетках лабильные структуры из микротрубочек, которые возникают и вновь распадаются.
Геометрию субъединиц внутри спиралей можно представить, построив проекцию развернутой поверхности структуры, как это показано на рис. 4-6, В. На приведенном рисунке субъединицы, образующие спираль, которая изображена на рис. 4-6, Б, лежат на плоскости, полученной мысленным разрезанием цилиндра вдоль оси и распрямлением цилиндрической поверхности. В рассматриваемом примере число субъединиц на один виток спирали составляет приблизительно 4,3, однако это число может быть и целым. Взаимодействия bk между субъединицами вдоль оси волокна иногда могут оказаться более сильными, чем взаимодействия типа aj. В этих случаях микротрубочка «разрывается» на концах за счет разрыва аj-связей. Если это явление имеет место в микротрубочках флагеллы эукариот, его можно непосредственно наблюдать при помощи электронного микроскопа.
На рис. 4-7 приведены красивые спиральные структуры четырех разных типов, образованные из отдельных молекулярных фрагментов. Это пиль Е. coli, нить актина (F-актин) из мышечного волокна, жгутик бактерии (Е. coli) и вирион вируса табачной мозаики. Считается, что каждая из этих структур состоит из большого числа протомеров одного типа. Наиболее детально изучена структура вирусной частицы. Известна, в частности, последовательность 158 аминокислотных остатков, образующих каждую из субъединиц вирусного белка (мол. вес = 17 500); число субъединиц на частицу равно примерно 2200, из них сформирована палочка длиной ~300 нм. В бороздке спирали располагается цепь РНК, содержащая приблизительно 6600 нуклеотидов (таким образом, на одну субъединицу белка приходится 3 нуклеотида) [34—36а]. Как и в более простой спиральной структуре пиля бактерии [37], белковые субъединицы вирусной частицы, по-видимому, связаны друг с другом только гетерологическими связями.
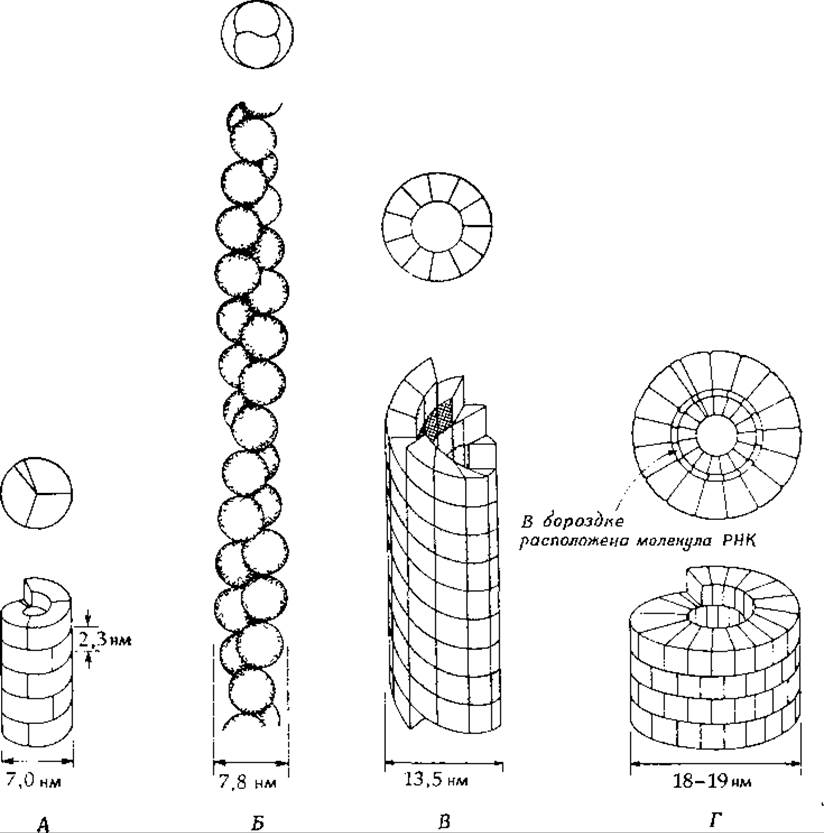
РИС. 4-7. Некоторые спиральные структуры, построенные из белковых субъединиц. А. Пиль Е. coli: шаг спирали 2,3 нм, диаметр отверстия 2,0—2,5 нм; мол. вес одной субъединицы 17 000; на виток спирали приходится ![]() субъединицы. Б. — F-актин мышцы: шаг спирали 70 им; две цепочки; мол. вес одной субъединицы 60 000; на один виток спирали приходится 13 субъединиц. В. Жгутик бактерии: шаг спирали 25 нм; диаметр отверстия 6 нм; пять цепочек; мол. вес одной субъединицы 40 000; на виток спирали приходится 11 субъединиц. Г. Вирус табачной мозаики: длина 300 нм (приблизительно 2200 субъединиц); шаг спирали 2,5 нм; мол. вес одной субъединицы 17 500; на виток спирали приходится 16,3 субъединицы.
субъединицы. Б. — F-актин мышцы: шаг спирали 70 им; две цепочки; мол. вес одной субъединицы 60 000; на один виток спирали приходится 13 субъединиц. В. Жгутик бактерии: шаг спирали 25 нм; диаметр отверстия 6 нм; пять цепочек; мол. вес одной субъединицы 40 000; на виток спирали приходится 11 субъединиц. Г. Вирус табачной мозаики: длина 300 нм (приблизительно 2200 субъединиц); шаг спирали 2,5 нм; мол. вес одной субъединицы 17 500; на виток спирали приходится 16,3 субъединицы.
Сравнительно недавно была детально изучена структура белковой оболочки нитевидного бактериофага Pf1 [38, 39]. Мол. вес белковых субъединиц этого бактериофага (дополнение 4-В) равен ~5000. Они имеют форму а-спиралей длиной 7 нм, образующих в составе бактериофага структуру типа левой спирали с шагом 1,5 нм, на один виток которой приходится 4,4 субъединицы (рис. 4-8). Белковые «палочки» расположены под углом к оси спирали и направлены внутрь спирали. Каждая палочка соприкасается с двумя другими, находящимися на 5 (—5) и на 9 (—9) единиц ниже по ходу спирали (рис. 4-8, А, Б). Такое расположение приводит к тому, что между субъединицами осуществляется гидрофобное взаимодействие типа ключ — замок (гл. 2, разд. Б.3,г) Весьма вероятно, что бактериальные пили и жгутики имеют именно такую структуру, а не структуру, изображенную на рис. 4-7.

РИС. 4-8. А. Предполагаемое расположение а-спиральных белковых палочек размером 1,0X7,0 нм в белковом чехле нитчатого бактериофага Рf1 [38]. Пунктирная линия в верхней части рисунка — левая а-спираль, на которой предположительно располагаются N-концы палочек. Б. Схематическое представление расположения а-атомов углерода в смежных а-спиральных палочках; атомы спроектированы на плоскость, параллельную оси вирусной частицы. Числа в кружочках — номера аминокислотных остатков. На примере трех субъединиц показано взаимное расположение одной из них (0) и находящихся ниже по ходу спирали субъединиц — пятой (—5) и девятой (-9) [39].
Как спиральные вирусы, так и пили бактерий могут рассматриваться как одиночные спирали из субъединиц, которые иногда называют спиралями с одним доступным концом. Актиновая нить (рис. 4-7) состоит из двух цепей, построенных из субъединиц; эти цепи закручены одна вокруг другой [40], т. е. структура имеет два доступных конца. Жгутики бактерий Е. coli и Salmonella (рис. 4-7, В) можно представить себе как пять цепочек, закрученных вместе вокруг одной и той же оси (одна из цепочек на рисунке для наглядности заштрихована). Те же жгутики можно, однако, рассматривать и как структуры, образованные 11 параллельными нитями, закрученными в спираль со значительно большим шагом [41]. Жгутики бактерий обладают многими удивительными свойствами (см. дополнение 4-Б). Так, например, на электронно-микроскопических фотографиях они обычно выглядят как «надспирали» с шагом, равным ~2,5 мкм. О каких особенностях молекулярной упаковки может свидетельствовать этот факт? Было бы очень хорошо, если бы читатель подумал и сам решил этот вопрос, но не слишком спешил с ответом.
Дополнение 4-А
Микротрубочки и действие колхицина
Важной составной частью цитоплазмы являются микротрубочки — полые стерженьки, наружный диаметр которых составляет 24±2 нм, а внутренний 13—15 нм. Наиболее удивительна их форма в жгутиках и ресничках эукариотических клеток (рис. 1-5). Устойчивые микротрубочки ресничек являются, по-видимому, неотъемлемой частью аппарата, обеспечивающего движение жгутикова,б. (Приведенный справа рисунок взят из работыб.) Лабильные (т. е. образующиеся, а затем распадающиеся) микротрубочки обнаруживаются чаще всего в цитоплазме клеток, способных к перемещению (например, в псевдоподиях амеб). Митотическое веретено (гл. 15, разд. Г.9) представляет собой набор микротрубочек, обеспечивающих перемещения хромосом в делящейся клетке. Микротрубочки обнаруживаются также в плоскостях деления растительных клеток.
Большое число микротрубочек содержится в длинных аксонах нервных клеток. Здесь они, вероятно, обеспечивают быстрый перенос белков и других веществ из тела клетки в аксон Микротрубочки, функция которых неизвестна, обнаружены и во многих сенсорных клетках. Недавно было показано, что микротрубочки содержатся в цитоплазме самых разных клеток. Используя непрямой метод флуоресцирующих антител, Вебер и др.в получили приведенную ниже (с любезного разрешения автора) микрофотографию фибробластов мышиного эмбриона.
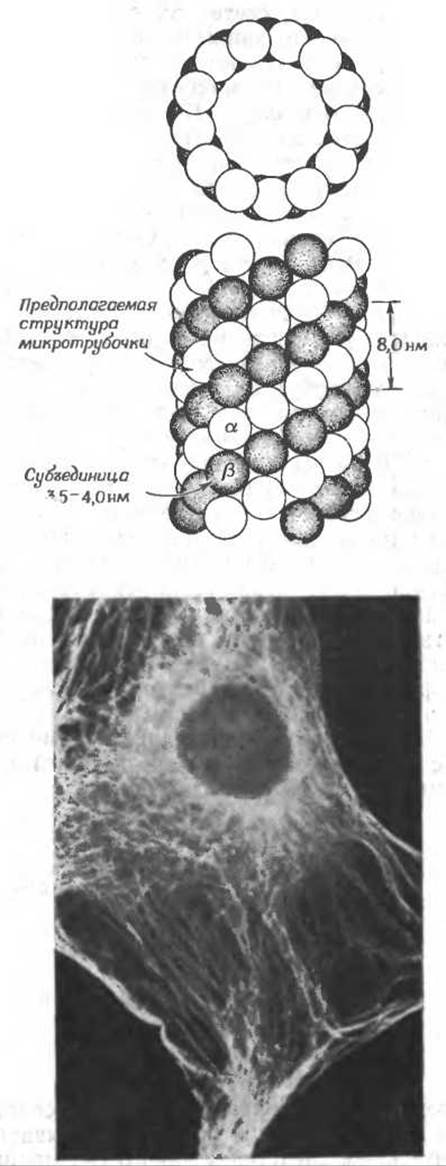
Клетки фиксировали формальдегидом, обезвоживали и подвергали воздействию антител, полученных путем иммунизации кроликов белком микротрубочек. Затем клетки обрабатывали флуоресцирующими антителами козы, специфичными в отношении кроличьего у-глобулина (дополнение 5-Е), и фотографировали в свете их собственной флуоресценции. Предполагается, что движения в системах, содержащих микротрубочки, осуществляются с участием других белков. Так, например, «ручки» на микротрубочках ресничек (рис. 1-5) катализируют гидролиз АТР, напоминая в этом отношении мышечный белок миозин. Движение ресничек осуществляется, вероятно, за счет скольжения микротрубочек по типу скольжения фибрилл в скелетной мышце (разд. Е).
Все микротрубочки построены из тубулинов — димеров, состоящих из близких по структуре субъединиц (а, ß) с мол. весом 60 000; в состав микротрубочек входит также в незначительном количестве белок с большим мол. весомг. Сами микротрубочки представляют собой, вероятно, группы параллельно расположенных нитей из соединенных друг с другом (конец в конец) молекул тубулина. Каждая димерная молекула тубулина связывает две молекулы GTP, причем одну из них прочнее. В этом отношении тубулин напоминает актин, субъединицы которого имеют приблизительно такие же размеры. Однако аминокислотные последовательности этих белков существенно различаются.
Обычно считается, что лабильные микротрубочки цитоплазмы находятся в динамическом равновесии с мономерными или димерными единицами. Известно, например, что микротрубочки могут образовываться или распадаться в зависимости от метаболических условий. Для их сборки нужен GTP, гидролиз которого, по-видимому, является существенной частью процесса сборкид. Недавно опубликованные данные о фосфорилировании белков микротрубочек указывают на то, что процесс этот очень сложен.
Особый интерес представляет взаимодействие микротрубочек с алкалоидом колхицином, который продуцируется различными членами семейства Liliaciae.

Это соединение, содержащее в своем составе трополоновое кольцо, специфически и прочно связывается с тубулином. Поразительным следствием такого связывания в живой клетке является разрушение лабильных микротрубочек, в том числе участвующих в образовании митотического веретена. При обработке делящихся клеток колхицином процесс деления задерживается на стадии метафазы (гл. 15, разд. Г.9), и образующиеся дочерние клетки обладают высокой степенью плоидности. Все это позволило широко использовать колхицин в качестве агента для получения тетраплоидных сортов цветущих растений. Аналогичное влияние на микротрубочки оказывают противоопухолевые агенты винкристин и винбластин — алкалоиды, синтезируемые малым барвинком (Vinca mіпоr)ж.
Микротрубочки ресничек (и жгутиков) эукариот имеют другую структуру. В большей части реснички они находятся в форме слившихся пар: А-трубочка содержит «ручку», и В-трубочка присоединяется к ней за счет общих субъединиц, расположенных в центре структуры. Как и из лабильных микротрубочек, было выделено два типа молекул тубулина, однако вопрос о том, как они вписываются в структуру спаренных микротрубочек, остается открытым.
a Snyder I. A., McIntosh I. R., Annu. Rev. Biochem., 45, 699—720 (1976).
б Bryan I., Fed. Proc., Fed. Am. Soc. Exp. Biol., 33, 152—157 (1974).
в Weber K., Pollack R., Bibring T., PNAS, 72, 459—463 (1975).
г Murphy D. B., Boris у G. G., PNAS, 72, 2696—2700 (1975).
д Jacobs M., Smith H., Taylor E. W., JMB, 89, 455—468 (1974).
e Marguhs T. N., JACS, 96, 899—902 (1974).
ж Wilson L., Bamberg J. R., Mizel S. B., Grisham L. M., Creswell К. M., Fed. Proc, Fed. Am. Soc. Exp. Biol., 33, 158—166 (1974).