Биохимия - Химические реакции в живой клетке Том 1 - Д. Мецлер 1980
Мембраны и клеточные оболочки
Строение клеточных стенок бактерий
Стенка грамотрицательных бактерий
С наружной стороны муреинового слоя у Е. coli и других грамотрицательных бактерий находится слой, содержащий фосфолипид, или наружная мембрана, сходный по толщине, а возможно, и по структуре с типичными биологическими мембранами (рис. 5-8). По-видимому, эта мембрана соединена с муреиновым слоем следующим образом. Небольшой гидрофобный белок, состоящий из 57 аминокислотных остатков, связывается с муреином амидной связью. Она образуется между аминогруппой боковой цепи С-концевого лизина белка и остатком диаминопимелиновой кислоты муреина [105]. Таким образом, белок заменяет один из концевых остатков D-аланина в среднем у одного из десяти пептидов муреина. Расположенный на N-конце белка серин в свою очередь ковалентно связывается с липидом, который, по-видимому, включен в состав наружной мембраны. На клетку приходится около 105 молекул гидрофобного белка, которые располагаются на поверхности пептидогликана площадью ~3 мкм2. Следовательно, расстояние между белковыми молекулами должно составлять в среднем ~5 нм.
У Е. coli между мембранами находится еще один, более крупный белок в количестве приблизительно 105 молекул на клетку. Молекулы этого белка оболочки (мол. вес ~ 36 500) образуют на внешней поверхности пептидогликана гексагональные структуры с расстоянием между молекулами 7,5 нм [106]. Возможно, белок оболочки придает жесткость палочковидным бактериям. В образовании упорядоченных структур на поверхности многих бактерий принимают участие и более крупные белки (мол. вес. 65 000—150 000)1 [106а].
Наружная поверхность внешней мембраны грамотрицательных бактерий покрыта удивительно сложно устроенным липополисахаридом [107, 108]. Внешний слой липополисахарида представляет собой совокупность длинных вытянутых полисахаридных цепочек, состоящих из повторяющихся специфических единиц, обладающих антигенными свойствами и получивших название О-антигенов. К этим полисахаридам могут быть получены специфические антитела. Структура полисахаридов характеризуется большим разнообразием — известно 1000 «серотипов» сальмонелл. Согласно существующей классификации, их разделяют на 17 основных групп. В группу Е3, например, входят серотипы, которые состоят из повторяющихся единиц
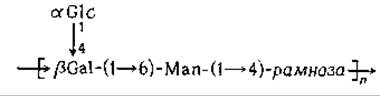
где число п в среднем равно ~50. Серотипы групп А, В и D содержат повторяющуюся единицу

где X — это 3,6-дидезоксигексоза : паратоза в группе А, абеквоза в группе В и тивелоза в группе D (рис. 5-10). Существование большого числа серотипов определяется изменчивостью компонентов, типами связей в повторяющихся единицах и структурными особенностями концевых участков цепей.
Часть О-антигена, обращенная к мембране, представляет собой более короткую полисахаридную цепочку, строение которой по сравнению с внешней частью характеризуется не столь сильным разнообразием. Однако в ее состав входят два сахара, обнаруживаемые только в стенках бактерий, а именно гептоза, содержащая 7 углеродных атомов, и а-кетосахарная кислота кетодезоксиоктонат, содержал 8 углеродных атомов. Структурные формулы этих сахаров приведены на рис. 5-10, а их расположение в липополисахаридах сальмонелл — на рис. 5-11. На последнем рисунке показано также, каким образом О-антиген соединяется с повторяющейся единицей, состоящей из двух молекул N-ацетилглюкозамина, связанных между собой ß-1,6-связью. Эти дисахаридные фрагменты соединены друг с другом при помощи пирофосфатных мостиков. Далее к свободным гидроксильным группам в дисахариде присоединены при помощи сложноэфирных связей остатки жирных кислот, которые, вероятно, проникают во внешнюю мембрану и играют роль якоря для всего липополисахарида. Соотношение жирных кислот — лауриновой, миристиновой и пальмитиновой — в дисахаридных единицах составляет 1:1:1. Кроме того, с дисахаридными единицами связаны три молекулы 3-D-оксимиристиновой кислоты. Два ее остатка связаны амидными связями с аминогруппами единиц GlcN, а третий образует за счет гидроксильной группы сложноэфирную связь с другой молекулой жирной кислоты. Считают, что именно эта специфическая структура ответственна за необычайно сильно выраженное пирогенное действие липополисахарида.
1 Кроме двух упомянутых выше белков в клеточной оболочке Е. coli имеется один белок, содержание которого сопоставимо с содержанием белка оболочки. См. обзор Гартеиа и др. [106b].

РИС. 5-10. Некоторые «редкие» сахара, входящие в состав «антигенов» наружной клеточной стенки грамотрицательных бактерий.
Почему у сальмонелл так много различных поверхностных антигенов? Ключом к ответу может служить тот факт, что концы описанных выше углеводных цепей являются именно теми группами, к которым присоединяются антитела животных при попадании бактерий в кровь. У ряда мутантов, известных под названием «R-формы» (от англ, rough — шероховатый, так как они образуют шероховатые колонии на агаре), внешний О-антиген отсутствует. R-мутанты сальмонеллы не патогенны, тогда как «гладкие» (не шероховатые) штаммы, содержащие интактный О-антиген, часто служат причиной болезни. Не исключено, что при определенном расположении сахарных колец на конце О-антигена организм хозяина не воспримет этот антиген как опасный. Так или иначе, ясно, что углеводные группы, расположенные на поверхностях клетки, играют важную роль в механизмах, при помощи которых клетки узнают друг друга.

РИС. 5-1). Повторяющаяся единица наружного гликолипидного слоя клеточной стенки Salmonella [108].
Интересные результаты были получены при исследовании умеренных бактериофагов — бактериальных вирусов, генетический материал которых может включаться в геном бактерий (гл. 15, разд. Г.8). Иногда включение вирусных генов в энтеробактерию вызывает изменение структуры О-антигена. Заражение одним вирусом приводит к потере О-ацетильных групп некоторыми сахарными остатками; другие вирусы вызывают появление дополнительных заместителей. Под влиянием ряда вирусов в определенных местах молекул олигосахаридов a-связи меняются на ß-связи или связи 1,4 на связи 1,6. Очевидно, вирусные гены могут, с одной стороны, кодировать синтез новых ферментов, гликозилтрансфераз, участвующих в построении О-антигенов, а с другой — подавлять синтез трансфераз хозяина. Интересно также, что патогенный микроорганизм — возбудитель брюшного тифа Salmonella typhi — представляет опасность лишь в том случае, если он заражен определенным умеренным бактериофагам. Здесь напрашивается явная аналогия с индукцией трансформации клеток млекопитающих в культуре онкогенными вирусами, при которой изменяются их поверхностные антигены.
Исследования умеренных фагов сальмонелл позволили понять некоторые особенности механизмов, с помощью которых эти бактериальные вирусы связываются со стенками клеток-хозяев. Местом первичного присоединения являются, по-видимому, сами О-антигены. Тонкие нити, расположенные на отростке фага (дополнение 4-Д), действуя на подобие антител, связываются со специфическими группировками полисахарида. Однако в результате включения генома фага и изменения строения О-антигена последующее присоединение вирусов блокируется. В то же время клетки бактерий становятся восприимчивыми к вирусам другого штамма [109].