Биохимия - Химические реакции в живой клетке Том 2 - Д. Мецлер 1980
Организация метаболизма: катаболические пути
Окисление жирных кислот
Кетоновые тела
При расщеплении жирной кислоты с четным числом атомов углерода путем ß-окисления последним промежуточным продуктом перед полным превращением в ацетил-СоА служит четырехуглеродный ацетоацетил-СоА:
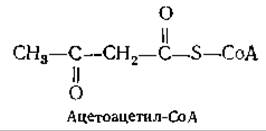
Ацетоацетил-СоА, который в организме, по-видимому, находится в равновесии с ацетил-СоА, является важным промежуточным продуктом [12а]. Он не только может расщепиться на две молекулы ацетил-СоА и таким образом войти в цикл трикарбоновых кислот, но служит также предшественником при синтезе изопреноидных соединений, в том числе и холестерина (гл. 12, разд. И). Не менее важное значение имеет ацетоацетат, один из компонентов крови.
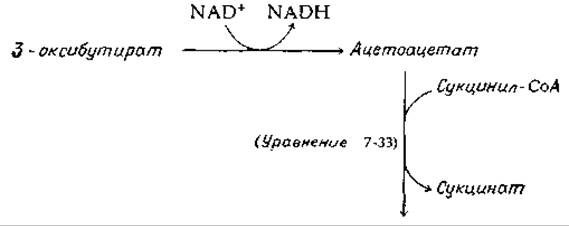
Ацетоацетат — это ß-кетокислота, которая может легко декарбоксилироваться в ацетон [уравнение (7-74)] и может также быть восстановлена NADH-зависимой дегидрогеназой в L-3-оксибутират. Отметим, что конфигурация этого соединения противоположна конфигурации D-3-оксибутирил-СоА, образующегося в ходе ß-окисления жирных кислот (рис. 9-1) и запасаемого в виде полимера (оксибутирата) в бактериях (гл. 2, разд. Д, 4).

Все три соединения, ацетоацетат, ацетон и 3-оксибутират, известны как кетоновые тела. Их концентрации сильно возрастают при различных патологических нарушениях, из которых наиболее известен сахарный диабет (дополнение П-В), а также при голодании. Развивающийся при этом кетоз при его сильных проявлениях очень опасен, так как образование кетоновых тел сопровождается освобождением ионов водорода [см. уравнение (9-7)] и закислением крови.
Кровь крысы в норме содержит около 0,07 мМ ацетоацетата, 0,18 мМ оксибутирата1) и варьирующее количество ацетона [13]. Ацетоацетат и оксибутират являются важными источниками энергии для мышечной и других тканей в тех случаях, когда в них поступает недостаточное количество углеводов. Ацетоацетат можно рассматривать как транспортную форму ацетильных остатков, которые могут вновь превращаться в ацетил-СоА и окисляться в цикле трикарбоновых кислот.
1) После 48 ч голодания эти концентрации возрастают до 0,5 и 1,6 мМ, в то время как концентрация глюкозы в крови (6 мМ) понижается до 4 мМ [13]
Небольшое количество свободного ацетоацетата образуется путем гидролиза ацетоацетил-СоА, однако большая его часть образуется в печени в результате двухстадийного процесса [уравнение (9-7)], который тесно связан с системой синтеза холестерина и других полипренильных соединений.

В уравнении (9-7) стадия а представляет собой альдольную конденсацию с последующим гидролизом одной тиоэфирной связи (гл. 7, разд. К, 2, е), тогда как стадия б является простым альдольным расщеплением. Стехиометрия полной реакции соответствует прямому гидролизу ацетоацетил-СоА до ацетоацетата. Вероятно, такой несколько усложненный механизм предоставляет больше возможностей для осуществления строгого контроля, однако не исключено, что это объясняется какими-то другими причинами.
Использование кетоновых тел в качестве источника энергии требует их обратного превращения в форму ацетил-СоА [уравнение (9-8)]. Кребс и др. [13] предположили, что в тканях, использующих энергию кетоновых тел, все реакции, представленные в уравнении 9-8, могут протекать в условиях, близких к равновесию.