БИОХИМИЯ - Л. Страйер - 1984
ТОМ 1
ЧАСТЬ I. КОНФОРМАЦИЯ И ДИНАМИКА
ГЛАВА 10. ВВЕДЕНИЕ В ПРОБЛЕМУ БИОЛОГИЧЕСКИХ МЕМБРАН
10.13. Липиды и многие мембранные белки быстро диффундируют в плоскости мембраны
Биологические мембраны-это не застывшие структуры. Напротив, и липиды, и многие белки мембран постоянно перемещаются в латеральном направлении. Быстрое движение белков мембраны выявляется с помощью флуоресцентной микроскопии при следующей постановке опыта. Культивируемые клетки человека и клетки мыши можно заставить слиться друг с другом; образующаяся при этом гибридная клетка называется гетерокарион. Одна часть плазматической мембраны гетерокариона происходит из клетки мыши, а другая - из клетки человека. Остаются ли мембранные белки мыши и человека разделенными в гетерокарионе или они смешиваются? Для ответа на этот вопрос использовали маркеры, а именно антитела с флуоресцентной меткой и далее визуально наблюдали за ними с помощью светового микроскопа. Антитело к мембранным белкам мыши имело зеленую флуоресценцию, а антитело к мембранным белкам человека-красную (рис. 10.26). В новообразованном гетерока- рионе одна половина поверхности светилась зеленым, а другая-красным. Однако меньше чем через час (при 37°С) участки зеленой и красной флуоресценции полностью смешивались. Этот опыт показывает, что мембранный белок способен диффундировать на расстояние порядка нескольких микрон примерно за 1 мин.
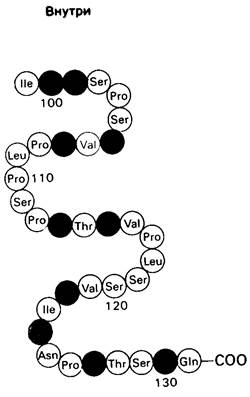
Рис. 10.26. Схематическое изображение слияния клетки мыши и клетки человека с последующей диффузией мембранных компонентов в плоскости мембраны. Спустя несколько часов после слияния маркеры с зеленой и красной флуоресценцией полностью перемешиваются
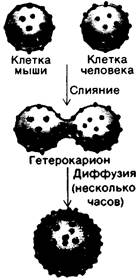
Более общий и количественный метод измерения латеральной диффузии мембранных компонентов в интактных клетках состоит в определении скорости восстановления флуоресценции после тушения (рис. 10.27). Для этого определенный компонент на поверхности клетки специфически метят флуоресцентным хромофором и помещают небольшую часть поверхности клетки (~ 3 мкм2) в поле зрения флуоресцентного микроскопа. Далее, находящиеся в поле зрения флуоресцирующие молекулы разрушают мощным пучком лазерных лучей. После этого определяют время нового появления флуоресценции в поле зрения, используя только слабое освещение, чтобы избежать дополнительного тушения. Если меченый компонент подвижен, то обесцвеченные молекулы исчезнут из поля зрения, а их место займут флуоресцирующие молекулы; в итоге интенсивность флуоресценции в поле зрения возрастет. Скорость восстановления исходного уровня флуоресценции зависит от латеральной подвижности флуоресцентно-меченого компонента, которую можно выразить через коэффициент диффузии О. Среднее расстояние 5 (см), пройденное по плоскости за время t (с), зависит от D (см2 • с-1) в соответствии с уравнением
s = (4 Dt)1/2.
Рис. 10.27. Метод восстановления флуоресценции после фототушения. А - флуоресценция меченого компонента клеточной поверхности на небольшом освещенном участке клетки. Б-флуоресцирующие молекулы обесцвечивают интенсивной световой вспышкой. В- по мере того как в результате диффузии обесцвеченные молекулы выходят из освещенной области, а не подвергшиеся обесцвечиванию входят в нее, происходит восстановление флуоресценции до исходного уровня. Г-скорость восстановления зависит от коэффициента диффузии
Коэффициент диффузии липидов в различных мембранах составляет примерно 10-8 см • с-1. Таким образом, молекула фосфолипида диффундирует в среднем на расстояние 2 • 10-4см, или 2 мкм, за 1 с. Это означает, что молекула липида может переместиться с одного конца бактериальной клетки на другой за 1 с. Экспериментально установленная величина коэффициента диффузии показывает, что вязкость мембран примерно в 100 раз выше вязкости воды и близка к вязкости оливкового масла.
В отличие от липидов белки очень неоднородны в отношении латеральной подвижности. Некоторые белки почти так же подвижны, как липиды, другие-практически неподвижны. Так, фоторецепторный белок родопсин характеризуется очень высокой подвижностью: его коэффициент диффузии равен 4 • 10-9 см2 с-1. С другой стороны, фибронектин (периферический гликопротеин, участвующий во взаимодействии клетки с субстратом) имеет коэффициент диффузии D менее 10-12 см с-1. Малая подвижность некоторых белков, возможно, объясняется тем, что они закреплены на субмембранных цитоплазматических структурах.
10.14. Мембранные белки не перемещаются поперек бислоев
В отличие от движения в плоскости мембраны спонтанное перемещение липидов от одной поверхности мембраны к другой происходит очень медленно. Перемещение молекулы с одной поверхности мембраны на другую называют поперечной диффузией (или «fliр-flор«-перескок), тогда как диффузию молекул в плоскости мембраны называют латеральной диффузией. Методом электронного парамагнитного резонанса было проведено прямое определение поперечной диффузии фосфолипидных молекул в фосфатидилхолиновых пузырьках; оказалось, что переход молекулы фосфолипида с одной стороны бислоя на другую совершается один раз за несколько часов (описание эксперимента дано в конце главы в разделе «Вопросы и задачи», пункт 5). Таким образом, поперечная диффузия молекулы фосфолипида на расстояние 50 А занимает в 109 раз больше времени, чем диффузия на то же расстояние в латеральном направлении.
Энергетический барьер для поперечной диффузии молекул белка еще выше, чем для липидов, поскольку в белках значительно больше полярных участков. Проведенные исследования не выявили поперечной диффузии белка. Следовательно, асимметрия мембран сохраняется на довольно длительное время.
10.15. Жидкостно-мозаичная модель биологических мембран
В 1972 г. Джонатан Сингер и Гарт Николсон (J. Singer, G. Nicolson) предложили жидкостно-мозаичную модель, объясняющую в общих чертах организацию биологических мембран. Согласно этой модели, мембраны представляют собой двумерные растворы определенным образом ориентированных глобулярных белков и липидов (рис. 10.28). В пользу предложенной модели свидетельствует большое количество экспериментальных данных. Основные положения жидкостно-мозаичной модели сводятся к следующему.
Рис. 10.28. Жидкостно-мозаичная модель

1. Большая часть мембранных фосфолипидов и гликолипидов представлена в виде бислоя. Липидный бислой играет двоякую роль, будучи одновременно растворителем для интегральных белков мембраны и барьером проницаемости.
2. Небольшая часть мембранных липидов специфически связана с определенными мембранными белками и, вероятно, необходима для их функционирования.
3. Мембранные белки свободно диффундируют в липидном матриксе в латеральном направлении, но не могут перемещаться в поперечном направлении, т. е. от одной поверхности мембраны к другой.