БИОХИМИЯ - Л. Страйер - 1984
ТОМ 1
ЧАСТЬ I. КОНФОРМАЦИЯ И ДИНАМИКА
ГЛАВА 10. ВВЕДЕНИЕ В ПРОБЛЕМУ БИОЛОГИЧЕСКИХ МЕМБРАН
10.16. Мембранам свойственна асимметрия
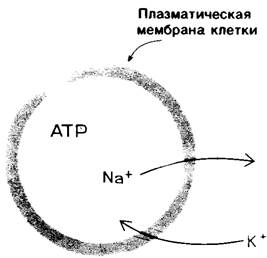
Мембраны асимметричны как по структуре, так и по функциям; об этом свидетельствуют примеры ориентации гликофорина и анионного канала, а также-более общий случай-локализация углеводов на наружной поверхности мембран. Наружная и внутренняя поверхности всех известных биологических мембран различаются по составу и ферментативной активности. Яркой иллюстрацией этого положения может служить насос, регулирующий концентрации Na+ и К+ в клетках. Эта транспортная система имеется в плазматических мембранах почти всех клеток высших организмов. Na+-K+-Hacoc ориентирован в плазматической мембране таким образом, что выводит Na+ из клетки и насасывает К+ в клетку (рис. 10.29). Для работы насоса требуется также АТР, который должен находиться на внутренней стороне мембраны. Специфический ингибитор насоса уабаин эффективен только при воздействии с наружной стороны мембраны.
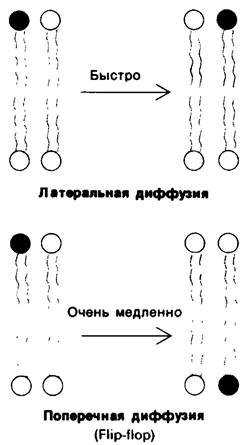
Рис. 10.29. Асимметрия системы сопряженного транспорта Nа+ и К + в плазматических мембранах
Как будет рассмотрено более подробно в одной из следующих глав (разд. 29.32), строго определенная ориентация мембранных белков обусловлена тем, что они синтезируются и включаются в мембрану, асимметричным образом. Эта абсолютная асимметрия сохраняется благодаря отсутствию трансмембранных перемещений белков на всем протяжении их существования в мембране. Липиды также распределены асимметрично, что обусловлено механизмом их биосинтеза, однако эта асимметрия практически во всех случаях, кроме гликолипидов, не является абсолютной. Например, в мембране эритроцита сфингомиелин и фосфатидилхолин расположены преимущественно в наружном слое бислоя, тогда как фосфатилэтаноламин и фосфатидилсерин главным образом-во внутреннем. Холестерол в большом количестве присутствует в обоих слоях бислоя. Функциональное значение асимметрии липидов пока еще не ясно.
10.17. Текучесть мембран зависит от состава жирных кислот и содержания холестерола
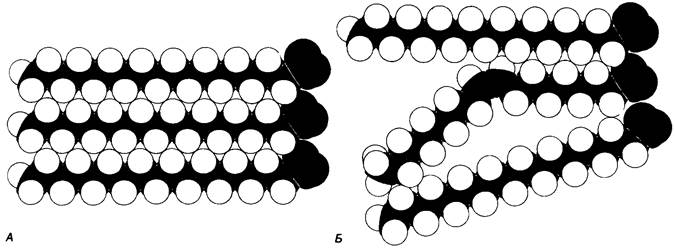
В мембранном бислое цепи жирных кислот в молекулах липидов могут находиться либо в строго упорядоченном жестком, либо в относительно дезорганизованном, жидком состоянии. В упорядоченном состоянии все связи С—С имеют транс-конформацию, тогда как в неупорядоченном -гош-конформацию (рис. 10.30). Переход от твердого (полностью транс-) к жидкому (частично гош-) состоянию происходит при повышении температуры выше точки плавления Тпл. Этот температурный переход зависит от длины цепи и степени ненасыщенности ацильного остатка. Наличие насыщенных ацильных остатков благоприятствует жесткому состоянию, так как прямые углеводородные цепи легко взаимодействуют между собой (рис. 10.31, А). Наличие же двойной связи цис-конфигурации приводит к изгибу углеводородной цепи, из-за которого нарушается строгая упорядоченность укладки ацильных остатков, и в результате Тпл снижается (рис. 10.31, Б). Температура перехода из жесткого состояния в жидкое зависит также от длины цепи. Длинные углеводородные цепи образуют более прочные связи друг с другом, чем короткие. В частности, каждая дополнительная группа —СН2— изменяет свободную энергию связи двух прилежащих углеводородных цепей на — 0,5 ккал/моль.
Рис. 10.30. Конформация связей С—С в углеводородных цепях остатков жирных кислот. А-транс (t)-конформация. Б и В- поворот на 120° дает гош (g)-конформацию, которая может быть g+ (поворот по часовой стрелке) или (поворот против часовой стрелки). При гош-конформации углеводородная цепь изгибается под углом 120°
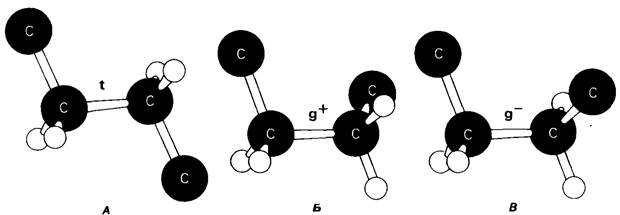
Рис. 10.31. Наличие двойных связей цис- конфигурации нарушает высокую степень упорядоченности упаковки углеводородных цепей жирных кислот. На приведенных пространственных моделях показана упаковка: А - трех молекул стеарата (С18, насыщенный), Б - молекулы олеата (С18, ненасыщенный) между двумя молекулами стеарата
Прокариоты регулируют текучесть своих мембран путем изменения числа двойных связей и длины ацильных цепей. Так, соотношение насыщенных и ненасыщенных остатков жирных кислот в мембране E. coli снижалось с 1,6 до 1,0 при понижении температуры среды с 42 до 27°С. Такое уменьшение доли насыщенных жирных кислот предотвращает чрезмерное затвердевание мембраны при пониженной температуре. У эукариот ключевым регулятором текучести мембран является также холестерол. Находясь между ацильными цепями, холестерол препятствует их кристаллизации. В сущности, из-за холестерола исчезает фазовый переход. С другой стороны, холестерол стерически блокирует сильное перемещение ацильных цепей и тем самым снижает текучесть мембран. Таким образом, благодаря этим взаимопротивоположным эффектам холестерола текучесть мембран поддерживается на каком-то среднем уровне.