БИОХИМИЯ - Л. Страйер - 1984
ТОМ 3
ЧАСТЬ V. МОЛЕКУЛЯРНАЯ ФИЗИОЛОГИЯ
ГЛАВА 35. ДЕЙСТВИЕ ГОРМОНОВ
35.17. Стероидные гормоны активируют специфические гены
Первичный эффект стероидных гормонов, в частности эстрадиола, прогестерона и кортизона, состоит в воздействии на выражение генов, а не непосредственно на активность фементов или процессы транспорта. В отличие от адреналина эти стероидные гормоны оказывают свое действие, только проникнув в клетку-мишень. Кроме того, местом их первичного действия служит не плазматическая мембрана, а клеточное ядро. Для полного проявления биологического эффекта стероидных гормонов требуются часы, а не минуты, поскольку он зависит от синтеза новых белков. Актиномицин D тормозит действие указанных стероидов, из чего следует, что оно связано с синтезом новой мРНК.
17Р-эстрадиол стимулирует увеличение матки. Первый этап этого воздействия состоит в том, что гормон связывается со специфическим рецептором в цитоплазме клеток матки. Связывание характеризуется высокой прочностью (К - 10-9M). Образовавшийся гормон-рецепторный комплекс мигрирует далее в клеточное ядро. В резуль тате связывания рецептора с эстрадиолом сродство рецептора к ДНК значительно повышается. Кроме того, у рецептора, связавшего зстрадиол, появляется вторая субъединица. Последнее находит отражение в увеличении коэффициента седиментации с 4S до 5S. Остается неизвестным, чем определяется специфичность участков ДНК, связывающих эстрадиольный рецептор: последовательностью ли оснований или белками хромосом. Взаимодействие гормон-рецепторного комплекса с ДНК высокоспецифично. Кроме того, строго специфична и активация рецептора путем связывания гормона. К рецептору эстрадиола может присоединиться эстрон, но этот комплекс не взаимодействует с ДНК. Неспособность эстрон-рецепторного комплекса к связыванию с ДНК вполне согласуется с тем, что эстрон не стимулирует увеличения матки.
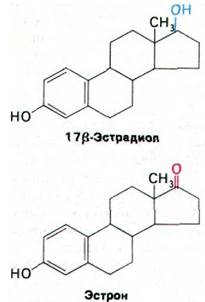
По-видимому, описанный механизм развития гормонального эффекта характерен не только для эстрадиола, но и в целом для всех стероидных гормонов. Так, в опытах с культурой ткани гепатомы было показано, что глюкокортикоидный стероидный гормон дексаметазон тоже связывается со специфическим цитоплазматическим рецептором; рецептор, связавший гормон, претерпевает конформационные изменения и затем мигрирует в клеточное ядро, где образует комплекс со специфическими участками в ДНК. Число участков ДНК, связывающих этот рецептор, определено равным 1 на 106 пар оснований. Полученное число согласуется с данными о том, что дексаметазон влияет на транскрипцию лишь небольшого числа генов. Каков механизм этой высокой избирательности в регуляции транскрипции со стороны стероидных гормонов, остается интригующей загадкой.
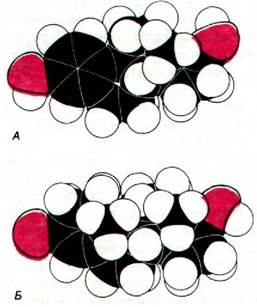
Рис. 35.22. Пространственные модели 17β-эстрадиола (А) и тестостерона (Б)
35.18. Белковые факторы роста типа ФРН и ЭФР стимулируют пролиферацию клеток-мишеней
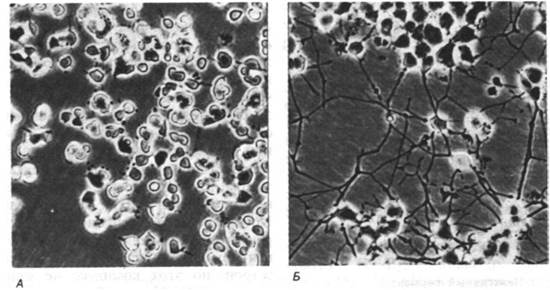
Как регулируется рост эукариотических клеток? Проведенное в последние годы выделение белковых факторов роста, способных специфическим образом стимулировать клетки-мишени, внесло существенный вклад в решение этого интересного и важного вопроса. Рита Леви-Монтальчини (Rita Levi-Montalcini) открыла, что фактор роста нервов (ФРН) играет решающую роль в развитии симпатических нейронов и определенных сенсорных нейронов у позвоночных. Под влиянием ФРН начинается деление и дифференцировка этих клеток. Чувствительным тестом на присутствие фактора роста нервов служит отрастание аксонов от культивируемых ганглиев (рис. 35.23). Биологически активная молекула ФРН состоит из двух идентичных полипептидных цепей массой по 13 кДа. Этот димер (называемый β-субъединицей) накапливается в месте своего синтеза - поджелудочной железе - в виде комплекса, имеющего субъединичную структуру α2γ2β и массу 130 кДа. γ-Субъединица является протеолитическим ферментом, а α-субъединица - ингибитором этой протеиназы. ФРН синтезируется в виде состоящего из α- и β-субъединиц прогормона, который в последующем расщепляется γ- протеиназой. По последовательности аминокислот ФРН напоминает инсулин. Следует отметить, что инсулин не только сильно активирует анаболические процессы в мышцах, печени и жировой ткани, но и оказывает стимулирующий эффект на рост большинства клеток. Можно предположить, что гены ФРН и инсулина произошли от общего предшественника.
Рис. 35.23. Фактор роста нервов (ФРН) индуцирует отрастание аксонов от культивируемых нервных клеток. А -без ФРН; Б - в присутствии ФРН
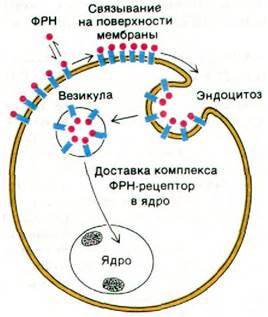
Был выделен и изучен также эпидермальный фактор роста (ЭФР). Это полипептид массой 6 кДа (рис. 35.24), способный стимулировать рост эпидермальных и эпителиальных клеток. ЭФР прочно связывается на плазматической мембране клеток- мишеней. Константа диссоциации комплекса ЭФР—рецептор составляет около 10-10 М. Спустя несколько минут после связывания ЭФР на мембране кластеры ЭФР-рецепторных комплексов попадают внутрь клетки путем эндоцитоза. Участками эндоцитоза являются так называемые окаймленные ямки (вдавления мембраны), содержащие клатрин (разд. 29.32). Далее пузырьки с ЭФР сливаются с лизосомами. Аналогичным образом попадают внутрь клеток и ФРН-рецепторные комплексы (рис. 35.25). Остается неизвестным, насколько необходим собственно процесс проникновения этих комплексов в клетку для передачи стимулирующего сигнала клеточному ядру.
Рис. 35.24. Последовательность аминокислот в эпидермальном факторе роста (ЭФР)
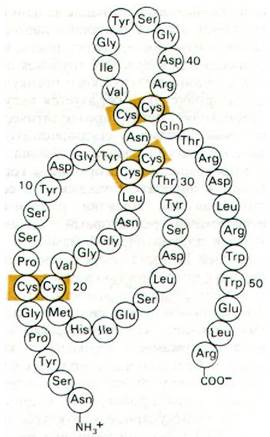
Рис. 35.25. Проникновение в клетку комплекса ФРН-рецептор