БИОХИМИЯ - Л. Страйер - 1984
ТОМ 3
Часть IV ИНФОРМАЦИЯ
ГЛАВА 26. ГЕНЕТИЧЕСКИЙ КОД Т ЗАВИСИМОСТЬ МЕЖДУ ГЕНАМИ И БЕЛКАМИ
26.7. Основные свойства генетического кода
Были расшифрованы все 64 кодона (табл. 26.4). 61 триплет соответствует определенным аминокислотам, а три кодируют терминацию. Поскольку существует 20 аминокислот и 61 триплет для их кодирования, очевидно, что код в высокой степени вырожден. Иными словами, многие аминокислоты детерминируются более чем одним триплетом. Только триптофан и метионин кодируются всего одним триплетом. Остальные 18 аминокислот кодируются двумя и более триплетами. Так, в частности, лейцин, аргинин и серин кодируются шестью кодонами каждый. В нормальных физиологических условиях код однозначен: каждый кодон обозначает только одну аминокислоту.
Таблица 26.4. Генетический код
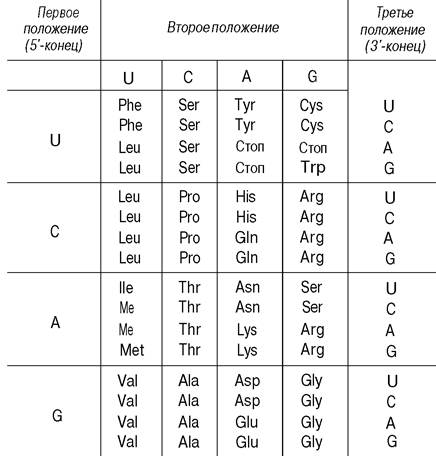
1 Зная положение оснований в кодоне, можно найти соответствующую аминокислоту. Например, кодон 5-AUG-3' в мРНК детерминирует метионин, тогда как CAU детерминирует гистидин. Кодоны UAA, UAG и UGA - сигналы терминации (стоп-кодоны). AUG является частью сигнала инициации помимо того, что он кодирует внутренние остатки метионина. В действительности оказалось, что только три кодона не кодируют никакой аминокислоты: UAG, UAA и UGA.
Кодоны, соответствующие одной аминокислоте, называют синонимами. Например, CAU и САС - синонимы для гистидина. Обратите внимание, что синонимы не разбросаны случайным образом по таблице генетического кода (табл. 26.4). Аминокислота, кодируемая двумя или более синонимами, занимает одну клетку в таблице (за исключением тех случаев, когда для данной аминокислоты существует более четырех синонимов). Аминокислоты, расположенные в одной клетке, кодируются кодонами, у которых два первых основания одинаковые, а третье различается, например, GUU, GUC, GUA и GUG. Большинство синонимов различается только последним основанием триплета. Рассмотрение кода показывает, что XYC и XYU всегда кодируют одну и ту же аминокислоту, a XYG и XYA чаще (но не всегда) кодируют одну и ту же аминокислоту. Структурные основы такой эквивалентности кодонов станут понятны после обсуждения природы антикодонов в молекулах тРНК (разд. 27.6).
Каков биологический смысл сильной вырожденности генетического кода? Один из возможных ответов состоит в том, что вырожденность сводит к минимуму пагубное действие мутаций. Если бы код не был вырожден, 20 кодонов кодировали бы аминокислоты, а 44 вызывали бы терминацию цепи.
Таким образом, вероятность превращения кодона в сигнал терминации была бы гораздо выше в случае невырожденного кода, чем в существующем коде. Важно учесть, что мутации, приводящие к образованию сигнала терминации цепи, обычно приводят к синтезу неактивных белков, тогда как замещение одной аминокислоты другой обычно относительно безвредно. Кроме того, вырожденность кода может иметь определенное значение постольку, поскольку она позволяет нуклеотидному составу ДНК меняться в широких пределах, не влияя на аминокислотную последовательность белков, кодируемых этой ДНК, ([G] + [С])-содержание бактериальных ДНК колеблется от 30% до более 70%. Молекулы ДНК с сильно различающимся содержанием [G] + [С] могут кодировать одни и те же белки благодаря систематическому использованию различных синонимов.
26.8. Сигналы инициации и терминации синтеза белка
Как мы уже упоминали, UAA, UAG и UGA (стоп-кодоны) обозначают терминацию цепи. Эти кодоны считываются не молекулами тРНК, а особыми белками - факторами терминации. Сигнал начала (инициации) синтеза белка более сложен. Полипептидные цепи у бактерий начинаются с модифицированной аминокислоты формилметионина (fMet).
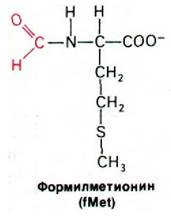
Существует особая тРНК, которая переносит fMet. Эта fMet-тРНК узнает кодон AUG (или реже GUG). Однако AUG является также кодоном для метионина, расположенного внутри полипептида, а GUG-кодон внутреннего валина. Это значит, что сигнал для первой аминокислоты полипептидной цепи должен быть сложнее, чем для всех последующих аминокислот. AUG (или GUG) представляет собой часть сигнала инициации. Существует другой сигнал, предшествующий AUG (или GUG), который определяет, будет ли кодон считан как сигнал инициации цепи или как кодон для внутреннего остатка метионина (или валина). Механизмы инициации и терминации синтеза белка мы рассмотрим в следующей главе (разд. 27.13).
26.9. Генетический код универсален
Как мы уже сказали, генетический код был расшифрован в результате исследований поведения тринуклеотидов и синтетических матричных РНК в бесклеточных системах, полученных из бактерий. Возникают два вопроса. Одинаков ли генетический код in vivo и in vitro? Одинаков ли генетический код у всех организмов? Убедительный ответ на эти вопросы был получен при анализе мутаций у вирусов, бактерий и высших организмов. Известно множество аминокислотных замен, возникающих в результате мутаций в генах гемоглобина человека, белка оболочки вируса табачной мозаики (ВТМ) и α-цепи триптофан-синтазы E. coli. Почти все эти аминокислотные замены объясняются заменами всего лишь одного основания (табл. 26.5). Это - надежная проверка правильности всего генетического кода и его универсальности.
Таблица 26.5. Мутации в гемоглобине человека, триптофан-синтазе E. coli и белке оболочки вируса табачной мозаики (ВТМ)

Почему код не изменился за миллионы лет эволюции? Обсудим действие мутации, изменяющей считывание мРНК. Такая мутация изменит последовательность аминокислот в большинстве, если не во всех белках, синтезируемых в клетках мутантного организма. Многие их этих изменений, безусловно, окажутся губительными, и, следовательно, должно существовать сильное давление отбора против таких мутаций.