БИОХИМИЯ - Л. Страйер - 1984
ТОМ 3
Часть IV ИНФОРМАЦИЯ
ГЛАВА 27. СИНТЕЗ БЕЛКА
27.11. Информационная РНК транслируется в направлении 5' → 3'
Направление считывания мРНК определяли с помощью синтетического полинуклеотида
![]()
использованного в качестве матрицы в бесклеточной системе синтеза белка. ААА кодирует лизин, а ААС-аспарагин. Полипептидный продукт имел структуру
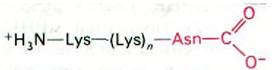
Поскольку аспарагин был расположен на карбоксильном конце, кодон ААС был считан последним. Следовательно, трансляция идет в направлении 5' → 3'. Напомним, что мРНК также синтезируется в направлении 5' → 3' (разд. 25.14). Это означает, что мРНК может транслироваться уже в то время, когда она еще синтезируется. Действительно, у E. coli 5'-конец мРНК взаимодействует с рибосомами вскоре после того, как он синтезируется (рис. 27.15). Таким образом, трансляция тесно сопряжена с транскрипцией.
Рис. 27.15. Транскрипция участка ДНК E. coli и трансляция новообразующейся мРНК. Транскрибируется лишь часть хромосомы

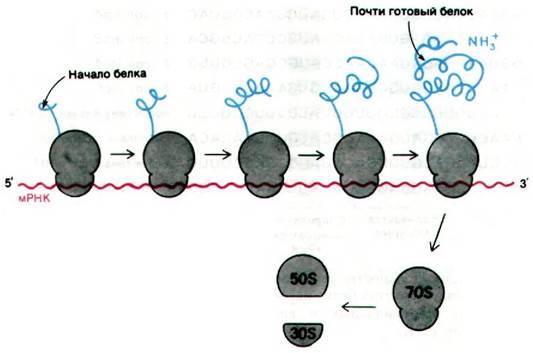
27.12. Одну молекулу мРНК одновременно транслирует несколько рибосом
Одну молекулу мРНК могут одновременно транслировать несколько рибосом. Это существенно увеличивает эффективность использования мРНК. Группа рибосом, связанных с одной молекулой мРНК, называется полирибосомой или полисомой. Рибосомы, входящие в состав этой структуры, работают независимо, и каждая из них синтезирует полную полипептидную цепь. При максимальной плотности рибосом на мРНК одна рибосома приходится на 80 нуклеотидов. Полирибосомы, синтезирующие гемоглобин (цепь гемоглобина содержит 145 аминокислот, а мРНК - соответственно 500 нуклеотидов), обычно состоят из пяти рибосом. Рибосомы, расположенные ближе всего к 5'-концу мРНК, несут самые короткие полипептидные цепи, а те, что ближе к 3'-концу, почти полные цепи. После освобождения полипептидного продукта рибосомы диссоциируют на 30S- и 50S-субчастицы.
27.13. Синтез белка в бактериях инициируется формилметиониновой тРНК
Как начинается синтез белка? A priori простейшая возможность состоит в том, что транслируется вся мРНК, и никакие особые сигналы начала синтеза не нужны. Однако эксперименты показали, что трансляция не начинается непосредственно на 5'-конце мРНК. На самом деле первый транслируемый кодон почти всегда расположен на расстоянии не менее чем 25 нуклеотидов от 5'-конца. К тому же многие молекулы мРНК у прокариот полицистронны, т.е. кодируют две или более полипептидные цепи. Например, одна молекула мРНК длиной около 7000 нуклеотидов кодирует пять ферментов пути биосинтеза триптофана у E. coli. Каждый из этих пяти ферментов имеет собственные сигналы начала и конца трансляции в мРНК. На деле все изученные молекулы бактериальных мРНК содержат сигналы начала и конца каждой кодируемой ими полипептидной цепи.
Рис. 27.16. Схематическое изображение организации полирибосомы (полисомы). Рибосомы движутся вдоль мРНК в направлении 5' → 3'. Рибосомы работают независимо друг от друга
Ключом к изучению механизма инициации цепи стало открытие того факта, что примерно половина N-концевых аминокислот белков E. coli - метионин, хотя в других положениях полипептидной цепи эта аминокислота встречается редко. Кроме того, N -конец новообразованных белков обычно модифицирован, из чего следует, что в инициации, очевидно, участвует какое-то производное метионина. Действительно, синтез белка у бактерий начинается с формилметионина (fMet). Существует особая тРНК, которая переносит формилметионин к рибосоме для инициации синтеза белка. Эта инициаторная тРНК (сокращенно обозначается mPHK¡) отличается от той тРНК, которая вводит метионин во внутренние положения (сокращенно тРНКm). Буква f указывает, что метионин, присоединенный к инициаторной тРНК, может быть формилирован, тогда как метионин, присоединенный к тРНКm, нет.
Рис. 27.17. Образование формилметио- нил-тРНК1
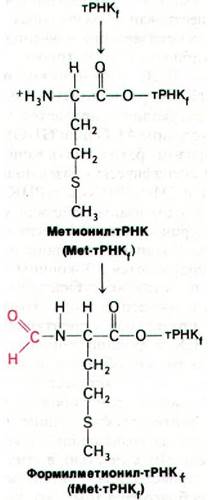
Метионин присоединяет к этим двум разновидностям тРНК одна и та же аминоа- цил-тРНК—синтетаза. Затем специальный фермент формилирует аминогруппу метионина, присоединенного к тPHKi. Донор активированной формильной группы в этой реакции N10-формилтетрагидрофолят.
27.14. Сигналом инициации служит кодон AUG (или GUG), которому предшествует несколько оснований, способных спариваться с 16S-PHK
Какие структурные особенности мРНК позволяют системе синтеза белка различать кодоны AUG? Первым шагом на пути к решению этого вопроса было выделение инициаторных участков ряда мРНК. Для этого комплексы мРНК с рибосомами, образовавшиеся в условиях, допускающих инициацию, но не элонгацию, расщепляли панкреатической рибонуклеазой. Во всех случаях от расщепления была защищена последовательность длиной около 30 нуклеотидов. Как и предполагалось, каждый из этих инициирующих участков содержал кодон AUG или GUG (рис. 27.18).
Рис. 27.18. Последовательности участков инициации синтеза белка некоторых бактериальных и вирусных РНК

Кроме того, каждый участок инициации содержит богатую пуринами последовательность, центр которой находится на расстоянии примерно 10 нуклеотидов в 5'-сторону от инициирующего кодона. Роль этого богатого пуринами участка стала ясна, когда была расшифрована последовательность 16S-PHK. 3'-конец этой рибосомной РНК содержит последовательность из нескольких оснований, комплементарную богатому пурином участку в областях инициации мРНК. Из продуктов ферментативного расщепления инициирующего комплекса удалось выделить комплекс 3'-концевого фрагмента 16S-мРНК и области инициации мРНК (рис. 27.19). Последовательности более чем 70 изученных участков инициации показывают, что число пар оснований, которые мРНК образует с 16S-PHK, колеблется от 3 до 9. Интересно, что изменение силы этого взаимодействия дает возможность регулировать эффективность инициации. Итак, место начала синтеза белка определяется взаимодействиями двух типов: спариванием оснований мРНК с 3-концом 16S-pPHKи с формилметиониновой инициаторной тРНК.
Рис. 27.19. Спаривание богатого пурином участка (показано синим цветом) в области инициации мРНК и 3'-концевого участка 16S-pPHK (красный цвет). Кодон AUG (зеленый цвет) определяет начало полипептидной цепи. Показанная на рисунке мРНК кодирует А-белок фага R17
