БИОХИМИЯ - Л. Страйер - 1984
ТОМ 3
Часть IV ИНФОРМАЦИЯ
ГЛАВА 24 ДНК: ГЕНЕТИЧЕСКАЯ РОЛЬ, СТРУКТУРА И РЕПЛИКАЦИЯ
24.5. Комплементарные цепи служат матрицами друг для друга при репликации ДНК
Модель двухспиральной молекулы ДНК позволила сразу же предложить механизм репликации ДНК. Уотсон и Крик опубликовали свою гипотезу через месяц после того, как представили модель структуры ДНК в виде прекрасной статьи, отличавшейся простотой и ясностью.
«Если задан определенный порядок оснований в одной из цепей, можно написать точную последовательность оснований в другой цепи, поскольку спаривание специфично. Таким образом, каждая из цепей комплементарна другой цепи, и именно это свойство подсказывает, каким образом может удваиваться молекула дезоксирибонуклеиновой кислоты.
Прежние дискуссии о самоудвоении всегда опирались на представление о некой матрице или каком-то шаблоне. Предполагалось, что матрица копирует непосредственно самое себя, или должна образовывать «негатив», который в свою очередь действует в качестве матрицы и образует исходный «позитив». Ни в одном случае не было предложено детального объяснения, как это могло бы происходить на уровне атомов и молекул.
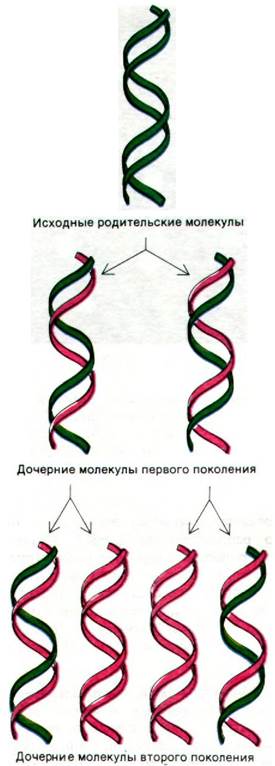
В нашей модели дезоксирибонуклеиновой кислоты имеется, по существу, пара матриц, причем каждая из них комплементарна другой. Мы полагаем, что перед удвоением водородные связи разрываются и две цепи раскручиваются и расходятся. Затем каждая цепь используется в качестве матрицы для образования на ней новой комплементарной цепи, так что в конце концов у нас будет две пары цепей, тогда как раньше была только одна. Более того, при таком способе репликация последовательность пар оснований будет в точности удвоена».
24.6. Репликация ДНК полуконсервативна
Уотсон и Крик предположили, что одна из цепей каждой дочерней молекулы ДНК синтезируется заново, а другая происходит от родительской молекулы ДНК. Такое распределение атомов родительской молекулы называют полуконсервативным. Метью Меселсон и Франклин Сталь (Matthew Meselson, Franklin Stahl) поставили принципиально важный эксперимент для проверки этой гипотезы. Родительскую ДНК пометили тяжелым изотопом азота 15N, чтобы сделать ее более тяжелой, чем обычная ДНК. Для этого выращивали Е. coli в течение многих поколений в среде, содержавшей в качестве единственного источника азота 15NH4Cl. Затем бактерии быстро переносили в среду, содержавшую 14N-стабильный изотоп азота. Этот эксперимент должен был показать, как распределяются изотопы 14N и 15N в молекулах ДНК в ходе последующих циклов репликации.
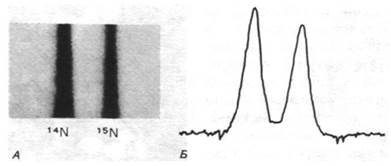
Распределение 14N и 15N в потомстве изучали с помощью только что разработанного метода равновесной седиментации в градиенте плотности. Небольшое количество ДНК растворяют в концентрированном растворе хлористого цезия с плотностью, близкой к плотности ДНК (~1,7 г/см3). Этот раствор центрифугируют почти до равновесного состояния. Под действием противодействующих процессов седиментации и диффузии в центрифужной ячейке возникает градиент концентрации хлористого цезия. В результате создается устойчивый градиент плотности от 1,66 до 1,76 г/см3. Под действием центробежной силы молекулы ДНК переходят в ту область градиента, в которой плотность раствора равна их собственной плавучей плотности. Высокомолекулярная ДНК образует четко выраженную полосу, обнаруживаемую по поглощению ультрафиолетового света. Молекулы 14N-ДНК и 15N-ДНК хорошо разделяются в таком градиенте, поскольку их плотность различается примерно на 1% (рис. 24.14).
Рис. 24.14. Разделение 14N- и 15N-ДНК при центрифугировании в градиенте плотности. А - фотография центрифужной ячейки в ультрафиолетовом свете; Б - денситометрическая кривая поглощения, полученная при сканировании фотографии, приведенной на рис. А
ДНК выделяли из бактерий через различные промежутки времени после переноса со среды, содержащей 15N, на 14N. Анализ этих образцов центрифугированием в градиенте плотности показал, что после одного цикла репликации ДНК дает одну полосу (рис. 24.15). Плотность этой полосы была равна в точности среднеарифметическому значению между плотностью 15N и 15N-ДНК. Отсутствие 15N-ДНК свидетельствовало о том, что целостность родительской ДНК в ходе репликации нарушается. Отсутствие 14N-ДНК показывало, что часть атомов всех дочерних молекул ДНК происходила от родительской ДНК. Соотношение 14N и 15N в дочерних молекулах должно составлять 1:1, так как плотность гибридных молекул ДНК была средней между 14N- и 15N-ДНК.
Рис. 24.15. Доказательство полуконсервативной репликации в клетках Е. coli с помощью центрифугирования в градиенте плотности. Положение полосы ДНК зависит от содержания 14N и 15N. После 1,0 генерации все молекулы ДНК представляют собой гибриды, содержащие равное количество 14N и 15N. Родительская ДНК (15N) после 1,0 генерации не обнаруживается
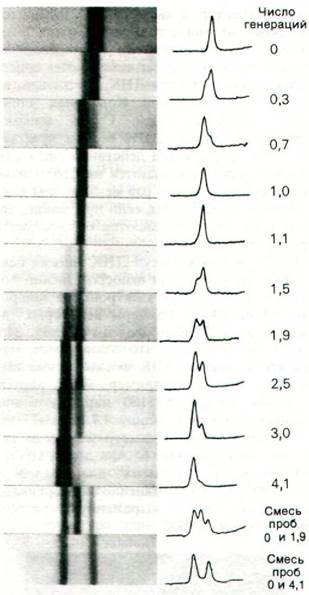
После двух циклов репликации ДНК распределялась поровну в двух полосах. Одна из них - гибридная ДНК, другая - 14N-ДНК. Меселсон и Сталь сделали из этих убедительных экспериментов вывод, «что азот молекул ДНК распределяется поровну между двумя физически раздельными структурами, что после удвоения каждая дочерняя молекула получает одну из этих структур и что эти структуры сохраняются интактными в течение многих удвоений». Полученные результаты прекрасно согласовывались с моделью репликации ДНК Уотсона- Крика (рис. 24.16).
Рис. 24.16. Схема полуконсервативной репликации. Родительская ДНК изображена зеленым, а новосинтезированная - красным