БИОХИМИЯ - Л. Страйер - 1984
ТОМ 3
ЧАСТЬ V. МОЛЕКУЛЯРНАЯ ФИЗИОЛОГИЯ
ГЛАВА 33. ИММУНОГЛОБУЛИНЫ
33.25. Разнообразие антител обусловлено соматической рекомбинацией многих генов клеток зародышевого пути и соматической мутацией
Подытожим теперь те механизмы, которые обеспечивают разнообразие антител. Клетки зародышевого пути содержат довольно большой набор генов вариабельных областей. Легкая цепь х кодируется несколькими сотнями (скажем, 300) генов V и пятью генами J. Существует по крайней мере три рамки, в которых происходит соединение генов V и J. Отсюда следует, что общее число полных генов Vx, образующихся при всех возможных сочетаниях генов V и J, составляет 300 • 5 • 3 = 4500. Подобным же путем может возникнуть аналогичное число вариантов тяжелых цепей. Соединение 4500 L-цепей с 4500 Н-цепями дает 4500 • 4500 = 2 • 107 различных по антигенной специфичности антител. Это число достаточно велико, чтобы обеспечить то большое разнообразие антител, которое образуется в организме животного.
Как упоминалось выше, генов Vӽ гораздо меньше, чем генов V. Так, у мышей имеется, по-видимому, только 2 гена V1. Однако соответствующих этому гену последовательностей аминокислот обнаружено намного больше. Представляется вероятным, что разнообразие легких цепей % возникает в результате соматических мутаций. В целом, как мы видим, природа использует каждый из трех обсуждавшихся выше (разд. 33.19) источников разнообразия - большой набор генов клеток зародышевого пути, соматические рекомбинации и соматические мутации: в итоге возникает тот богатейший набор антител, который необходим для защиты организма от вторжения чужеродных веществ.
33.26. Клонально-селекционная теория образования антител
В 50-х годах Нилс Ерне, Макферлейн Бёр- нет, Дэвид Тэлмедж и Джошуа Ледерберг (Niels Jerne, Macfarlane Burnet, David Talmage, Joshua Lederberg) разработали клонально-селекционную теорию, дающую обобщенное описание иммунного ответа.
Основные положения этой ныне общепринятой теории следующие.
1. Клетка, продуцирующая антитела, содержит ДНК с уникальной последовательностью оснований, определяющей последовательность аминокислот, характерную для синтезируемых ею цепей иммуноглобулина. Специфичность антитела целиком определяется последовательностью аминокислот.
2. Каждая отдельная клетка продуцирует антитела только одной специфичности. Следовательно, способность к синтезу определенного антитела заложена в клетке еще до контакта с антигеном.
3. Начинающая созревать клетка продуцирует небольшие количества антител одной специфичности. Часть из них прикрепляется к поверхности клетки. Если такая незрелая клетка встречается с антигеном, против которого оказались направлены ее антитела, то она погибает. Вследствие этого животное обычно не производит антител против собственных макромолекул, т. е. оно обладает толерантностью к своему. Действительно, клетки, продуцирующие антитела против собственных антигенов, элиминируются на протяжении эмбрионального развития. На зрелые клетки в отличие от незрелых встреча с антигеном действует стимулирующим образом: усиливается синтез антител и запускается клеточное деление. Потомки одной клетки составляют клон, обладающий всеми генетическими особенностями исходной клетки. Следовательно, деление клетки, инициированное контактом с антигеном, приводит к появлению клона клеток, синтезирующих антитела той же специфичности.
4. Клон проявляет тенденцию к сохранению и после исчезновения антигена. В случае нового появления того же антигена происходит стимуляция этих клеток, что и обусловливает иммунологическую память.
33.27. На поверхности клеток, продуцирующих антитела, имеются рецепторы антигенов
Молекулы специфических антител расположены на поверхности продуцирующих их клеток. Клетками-предшественниками являются лимфоциты. В обычных условиях они делятся медленно. Однако в результате стимуляции антигеном лимфоцит превращается в плазматическую клетку, очень активно синтезирующую и продуцирующую антитела. Если популяцию лимфоцитов пропустить через колонку, содержащую ковалентно присоединенные динитрофенильные (ДНФ) группы, то на колонке свяжется только очень небольшая часть клеток, а именно те, которые синтезируют антитела против ДНФ и несут молекулы таких антител на своей поверхности. Не задержавшиеся на колонке лимфоциты таких антител против ДНФ не вырабатывают.
Как происходит стимуляция лимфоцитов под действием специфических антител? В отсутствие антигена молекулы антител на поверхности лимфоцита распределены случайным образом. Добавление антигена оказывает поразительный эффект: расположенные на поверхности лимфоцита антитела вместе с присоединенными антигенами собираются вместе на одном конце клетки. образуя так называемый «колпачок» («кэп»). По завершении перераспределения молекулы антител оказываются внутри клетки. Образование «колпачка» идет с потреблением энергии и при участии сократительных элементов клетки. Итак, формирование на поверхности клетки решетки из комплексов антиген-антитело ведет к образованию «колпачка», а это стимулирует клеточное деление. Антиген должен быть поливалентным (т. е. содержать более одной специфической детерминанты) для того, чтобы создать перекрестные связи между молекулами антител на поверхности лимфоцита. Вопрос о зависимости митотической активности клеточного ядра от этих событий, разыгрывающихся на поверхности клетки, составляет интересную и важную область исследований.
Рис. 33.35. Полученное в сканирующем электронном микроскопе изображение бараньих эритроцитов, связавшихся с человеческим Т-лимфоцитом (в центре). Поверхность этого лимфоцита имеет рецепторы, специфичные в отношении компонентов бараньих эритроцитов
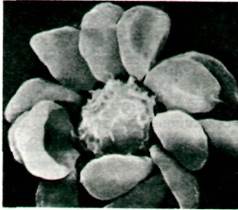
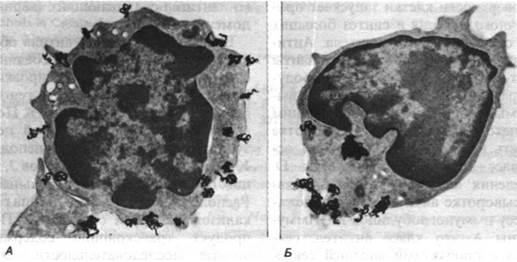
Рис. 33.36. Полученное методом радиоавтографии изображение иммуноглобулинов, образовавших «колпачок» на поверхности В-лимфоцита. Темные включения - меченные I антитела против иммуноглобулинов, добавленные к В-лимфоцитам из селезенки мыши. А - при 4°С молекулы иммуноглобулинов равномерно распределяются на поверхности клетки; Б - после инкубации при 37°С метка концентрируется в виде «колпачка» на одном полюсе клетки и затем попадает внутрь посредством эндоцитоза
33.28. Биологическое значение клональной селекции
Сущность теории селекции клонов в формулировке Бёрнета состоит в следующем: Ни один участок связывания антигена не адаптирован в эволюционном смысле в какой-либо определенной антигенной детерминанте. Структура участка, связывающего антиген, существует как таковая, и если она оказывается подходящей в том смысле, что сродство к данной антигенной детерминанте превышает какой-то определенный уровень, то инициируется иммунологически значимая реакция. Отбор предсуществующих вариантов, возникающих случайным образом, это не новая тема в биологии. В самом деле, в этом суть теории Дарвина. Любопытно отметить, что инструктивные теории предшествуют селекционным теориям: теория Ламарка предшествовала теории Дарвина, инструктивная теория в иммунологии была сформулирована раньше, чем клонально-селекционная теория.
Ерне высказал предположение, что механизмы отбора могут играть роль в функционировании нервной системы, например, в механизмах памяти. По существу, инструктивная и селекционная теории обучения были сформулированы очень давно. Локк (Lock) считал, что мозг человека подобен чистому листу бумаги, на котором опыт выписывает почти бесконечные узоры. Сократ, напротив, утверждал, что «все обучение - это напоминание о том, что уже существует в мозгу». Будем надеяться, что эта глава вам кажется чем-то давно знакомым!