БИОХИМИЯ - Л. Страйер - 1984
ТОМ 3
ЧАСТЬ V. МОЛЕКУЛЯРНАЯ ФИЗИОЛОГИЯ
ГЛАВА 33. ИММУНОГЛОБУЛИНЫ
33.22. Соединение генов V и I в различных рамках также способствует разнообразию антител
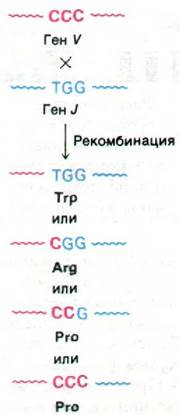
Второе удивительное открытие состояло в том, что набор из пяти генов J обеспечивал синтез не пяти, а большего числа последовательностей аминокислот для соответствующих областей легкой цепи. Анализ последовательностей аминокислот и оснований показал, что рекомбинация генов V и J происходит не абсолютно точно. Как оказалось, рекомбинация этих генов может иметь место по тому или иному из оснований вблизи кодона, детерминирующего остаток 95 (рис. 33.26). Следовательно, различие рамок, в которых происходит сращивание генов V и J, вносит дополнительный вклад в разнообразие антител в организме. Похоже, что иммунная система получает удовольствие от мелких погрешностей!
Рис. 33.26. Неточность места соединения генов V и J служит еще одним источником разнообразия антител
В каждом данном лимфоците экспрессирован только один из двух аллельных генов. Следовательно, все участки, связывающие антиген, продуцируемые отдельной клеткой, одинаковы. Обнаружена структурная основа такого избирательного выражения гена (называемого аллельным исключением). Как показал анализ фрагментов, полученных после рестрикции, неполный ген V правильно соединяется с геном J только в одной из двух гомологичных хромосом; экспрессируется же только правильно рекомбинированный ген иммуноглобулина.
33.23. мРНК для L- и Н-цепей образуются путем сращивания (сплайсинга) первичных продуктов транскрипции
мРНК легких ӽ-цепей содержит около 1250 оснований (рис. 33.27). Как и в других мРНК эукариот, в ней содержатся роlу(А)- последовательность, присоединенная к 3'-концу, и нетранслируемые последовательности на 3'- и 5'-концах. Лидерная последовательность вблизи 5'-конца мРНК х-цепи кодирует гидрофобную N-концевую область синтезируемой х-цепи. Образующаяся сигнальная последовательность (рис. 33.28) направляет рибосому к эндоплазматическому ретикулуму и определяет способность новосинтезированной полипептидной цепи иммуноглобулина проходить через мембрану ЭР в просвет его трубочек (разд. 29.30). Далее на внутренней стороне мембраны ЭР происходит отщепление сигнальной последовательности под действием пептидазы. Вариабельная и константная области легкой цепи кодируются смежной областью мРНК, которая следует непосредственно за лидерной последовательностью.
Рис. 33.27. Структура мРНК, кодирующей L-цепь

Рис. 33.28. Сигнальная последовательность новосинтезированной L-цепи. Желтым цветом отмечены гидрофобные остатки

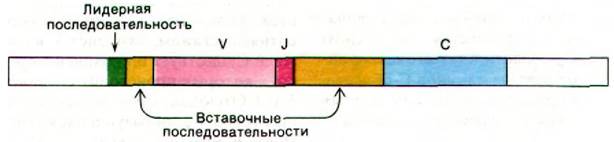
Первичный продукт транскрипции, из которого образуется эта мРНК, содержит две вставочные последовательности (рис. 33.29). Одна из них отделяет лидерную последовательность от начала мРНК, кодирующей вариабельную область, а вторая расположена между дистальным концом последовательности, комплементарным гену J, и началом последовательности, комплементарной гену С (т.е. кодирующей константный участок). При превращении первичного продукта в мРНК (процессинг) происходит удаление этих вставочных последовательностей. Любопытно, что ген J несет информацию для сплайсинга (сращивания) двух типов: одно - на уровне ДНК (слияние генов V и J) и другую - на уровне РНК (соединение J- и С-участков).
Рис. 33.29. Первичный транскрипт - предшественник мРНК L-цепи
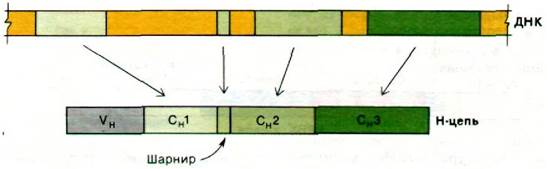
Какова структура гена, кодирующего тяжелую цепь? Вспомним, что Н-цепь состоит из четырех доменов: VH, СH1, СH2 и CH3. Недавно было осуществлено клонирование фрагмента ДНК, определяющего константную область тяжелой цепи IgG. Электронно-микроскопические исследования показали, что СH1, СH2 и СH3 кодируются разными участками ДНК (рис. 33.30). Еще один участок ДНК кодирует шарнирное соединение между СH1 и СH2. В целом доменная структура иммуноглобулинов (разд. 33.14) является отражением архитектуры соответствующих генов. Благодаря сплайсингу организмы оказались спо
собными создавать в ходе эволюции новые белки путем объединения участков ДНК, кодирующих разные домены.
Рис. 33.30. Три домена константной (С) области тяжелой цепи и шарнирный участок кодируются разными участками гена
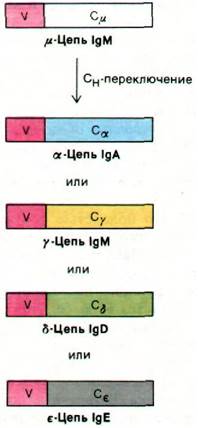
33.24. Разные классы антител образуются в результате перескока генов VH
Как уже упоминалось, существует пять классов иммуноглобулинов. Клетки, продуцирующие антитела, сначала продуцируют IgM, а затем IgG, IgA, IgD или IgE той же самой специфичности. При этом переключении от IgM на другой класс иммуноглобулинов легкая цепь остается неизменной. Более того, неизменной остается и вариабельная область тяжелой цепи. Меняется только константная область тяжелой цепи, и потому этот этап дифференцировки клетки, продуцирующей антитела, называют Сн-переключением (рис. 33.31).
Рис. 33.31. Синтез различных классов иммуноглобулинов. В результате того, что ген VH-области сначала соединяется с геном Сμ-области, а затем с каким-либо другим из генов С-области, формируются Н-цепи разных классов иммуноглобулинов
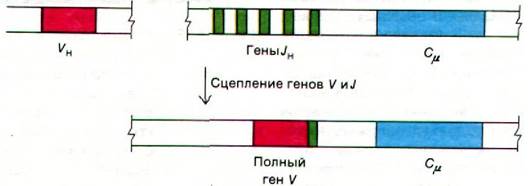
В эмбриональных клетках мыши гены, кодирующие константные области μ-, γ- и α- тяжелых цепей (и обозначаемых соответственно Сμ, Cγ и Cα, расположены в ряд, один за другим(рис. 33.32). Как оказалось, существует четыре гена константных участков γ-цепей, что полностью соответствует данным генетического анализа, выявившего четыре подкласса IgG. Рядом с геном Сμ локализуется набор расположенных тандемно генов J, кодирующих последний гипервариабельный участок вариабельной области. Полный ген тяжелой цепи IgM образуются путем транслокации гена VH к гену JH (рис. 33.33). В результате этой транслокации гены VH, JH и Сμ соединяются в функционально единый ген. Вставочные последовательности между лидерным отрезком и началом гена вариабельной области, между концом гена JH и началом гена Сμ, а также в пределах гена Сμ выстригаются в ходе превращения первичного траскрипта в мРНК для μ-цепи.
Рис. 33.32. Гены константных областей μ-, γ- и α-цепей расположены в ряд, один за другим. Положение генов Сδ и Сε еще не установлено
![]()
Рис. 33.33. В результате соединения генов Vн и JH образуется ген, кодирующий μ-цепь
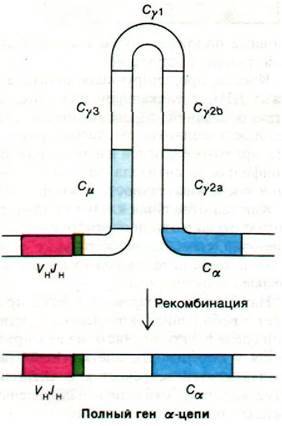
Как показал анализ продуктов рестриктазного расщепления ДНК из эмбриональных и миеломных клеток, ^-переключение происходит на уровне ДНК, а не РНК. Так, при переключении от IgМ к IgA ген VHJH, расположенный рядом с геном Сμ, перемещается в участок, расположенный рядом с геном Са (рис. 33.34). При этой рекомбинации гены между Сμ и Саобразуют петлю и выстригаются. Не исключено, что участки ДНК, претерпевающие рекомбинацию, несут палиндромную последовательность. Именно транслокация всего гена VHJHлежит в основе того факта, что IgA, продуцируемый определенной клеткой, идентичен по антигенной специфичности IgM, синтезированному той же клеткой на более ранней стадии развития. Каким образом клетка выбирает для транслокации один из нескольких генов Сн, остается неизвестным. Биологическое значение СH-переключения состоит в том, что весь домен, ответственный за узнавание (вариабельный домен), перемещается от первоначальной константной области (Сμ) к другим константным областям, кодирующим полипептидные цепи с иными эффекторными функциями.
Рис. 33.34. Структурная основа CH-переключения. В результате внутрихромосомной рекомбинации ген VHJH, находившийся возле гена Сμ, оказывается рядом с геном Са