БИОХИМИЯ - Л. Страйер - 1984
ТОМ 3
ЧАСТЬ V. МОЛЕКУЛЯРНАЯ ФИЗИОЛОГИЯ
ГЛАВА 33. ИММУНОГЛОБУЛИНЫ
33.18. Вариабельные и константные области кодируются разными, но соединившимися генами
Как организованы и как функционируют гены иммуноглобулина? Обнаружение четкого различия между вариабельными и константными областями в L- и Н-цепях дало основание думать, что гены иммуноглобулинов, так же, как и кодируемые ими полипептиды, имеют необычную архитектуру. Ранее уже упоминалось, что константные области х-цепей идентичны по всем положениям, кроме 191, где можег стоять либо лейцин, либо валин. Как показал анализ родословной, указанные два варианта (аллотипа) наследуются в соответствии с классическим правилом Менделя, а это служит веским доводом в пользу того, что константная область х-цепей кодируется только одним геном. Что касается вариабельных областей, то по данным генетического анализа они кодируются множеством генов. В 1965 г. Уильям Дрейер и Клод Беннетт (William Dreyer, Claude Bennett) выдвинули предположение, что в половых клетках множественные гены V пространственно отделены от одиночного гена С. Согласно этой гипотезе, в процессе дифференцировки клетки, продуцирующей антитело, один из генов V соединяется с геном С. Позднее стало известно, что действительно возможно сращивание ДНК (сплайсинг), аналогичное тому, которое имеет место, например, при включении ДНК фага в геном.
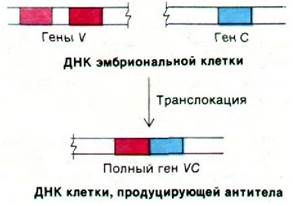
Окончательная проверка этой гипотезы транслокации сможет быть произведена только после того, как будет выделена чистая мРНК иммуноглобулина и будут разработаны методы анализа сложных геномов млекопитающих. Особенно интересно в этом отношении использование рестриктаз, расщепляющих большие хромосомы на специфические фрагменты ДНК, которые нетрудно далее подвергнуть анализу. Распределение генов V и С в этих фрагментах ДНК можно определить путем гибридизации их с фрагментами мРНК, специфичными в отношении либо вариабельной, либо константной области. В 1976 г. Сусуму Тонегава (Susumu Tonegawa), используя этот подход, определил, что в ДНК половых клеток гены V и С расположены далеко друг от друга, а в клетках, продуцирующих антитела, они тесно связаны. Следовательно, в ходе дифференцировки лимфоцитов происходит перемещение генов иммуноглобулина (рис. 33.23).
Рис. 33.23. При дифференцировке клеток, продуцирующих антитела, ген V транслоцируется, соединяясь с геном С
33.19. Как возникает разнообразие специфичности антител?
Организм животного обладает способностью к синтезу больших количеств специфических антител спустя несколько недель после введения практически любой чужеродной детерминанты. Число различных видов антител, которые могут синтезироваться в организме животного, огромно - вероятно, оно превышает миллион. Как мы уже видели, структурной основой специфичности антител служат последовательности аминокислот в вариабельных областях легких и тяжелых цепей. Это подводит нас к ключевому вопросу: каким образом возникают различные последовательности аминокислот вариабельных областей? Можно сформулировать в связи с этим и более конкретные вопросы.
1. Когда возникает разнообразие? На протяжении жизни животного (соматически) или на протяжении эволюции (генетически)?
2. Как возникает разнообразие? В результате случайных мутаций в ходе эволюции, путем соматической рекомбинации или путем соматической гипермутации?
Обилие данных относительно последовательностей аминокислот миеломных иммуноглобулинов позволило иммунологам разработать несколько гипотез относительно того, как возникло разнообразие антител. Были предложены три возможных механизма.
1. Гипотеза клеток зародышевых линий. Согласно этой модели, разнообразие свойственно уже клеткам зародышевых линий (и эмбриональным клеткам), содержащим
очень большое число (>104) генов, кодирующих вариабельные области антител. Каждой уникальной последовательности вариабельных областей соответствует участок ДНК в клетках зародышевого пути. Таким образом, эта гипотеза предполагает, что разнообразие возникло на протяжении эволюции в результате случайных мутаций и отбора.
2. Гипотеза соматических рекомбинаций. Согласно этой гипотезе, существует только небольшое число (порядка 100) генов, кодирующих вариабельные области антител. Эти гены, сходные между собой, но не идентичные, на протяжении жизни индивидуума претерпевают многократные рекомбинации в клетках, продуцирующих антитела. Предполагается, что кроссинговер идет внутрихромосомно. Расчеты показали, что рекомбинация 10 генов может привести к появлению 106 разных последовательностей аминокислот.
3. Гипотеза соматических гипермутаций. Согласно этой модели, разнообразие возникает в результате точковых мутаций одного-единственного гена вариабельной области данного подкласса. Однако только необычайно высокая частота мутаций могла бы создать на протяжении жизни индивидуума все разнообразие специфичности антител. Было высказано предположение, что в клетках, продуцирующих антитела (или в соответствующих клетках-предше- ственниках), увеличение скорости мутаций обеспечивается механизмом репарации ДНК, действие которого в данном случае направлено на производство ошибок.
33.20. Вариабельные участки L- и Н-цепей кодируются несколькими сотнями генов
Революция в методах изучения ДНК, произошедшая в последние годы, сделала вопрос о происхождении разнообразия антител доступным для экспериментального анализа. Особенно ценными в данном случае оказались методы клонирования генов и быстрого определения последовательности протяженных участков ДНК. Благодаря использованию этих методов за короткий срок была получена информация, необходимая для ответа на два узловых вопроса: сколько генов вариабельных областей содержится в клетках зародышевого пути? Меняется ли последовательность оснований в процессе дифференцировки клеток, продуцирующих антитела?
Ответ на первый вопрос сводится к тому, что существует несколько сотен генов вариабельных областей х-легких (Vz) и тяжелых цепей (VH). 300 генов Vx и 300 генов Vн, соединяясь в различных комбинациях (300 х 300), способны были бы кодировать 9 • 104 вариантов молекул антител, различающихся по специфичности. Этот расчетный максимум (9 • 104) намного ниже реального числа антител различной специфичности, синтезирующихся в организме животного; считается, что число таких антител значительно превышает 106. Расхождение между расчетом и реальной величиной еще выше в отношении легких цепей ![]() , которые, как было показано, кодируются менее, чем 10 генами V,, В целом число генов вариабельных областей в клетках зародышевого пути оказалось слишком малым, чтобы полностью обеспечить все разнообразие антител. Очевидно, в ходе дифференцировки лимфоцитов на протяжении жизни животного появляются какие-то дополнительные факторы, повышающие степень разнообразия.
, которые, как было показано, кодируются менее, чем 10 генами V,, В целом число генов вариабельных областей в клетках зародышевого пути оказалось слишком малым, чтобы полностью обеспечить все разнообразие антител. Очевидно, в ходе дифференцировки лимфоцитов на протяжении жизни животного появляются какие-то дополнительные факторы, повышающие степень разнообразия.
33.21. Открытие генов I (соединяющих) - дополнительного источника разнообразия антител
Следующий шаг в изучении механизма, обеспечивающего разнообразие антител, был сделан при определении последовательностей оснований клонированных генов, кодирующих иммуноглобулины в эмбриональных и миеломных клетках. Эти исключительно продуктивные исследования были проведены Тонегавой, Филиппом Ледером и Лероем Худом (Tonegawa, Philip Leder. Leroy Hood). Прежде всего, совершенно неожиданно выяснилось, чем в эмбриональных клетках гены V кодируют вариабельные области L- и Н-цепей вовсе не целиком. Ген V эмбриональных клеток (и клеток зародышевого пути) заканчивается кодом для аминокислотного остатка 95, а не 108, который составляет конец вариабельной области полипептидной цепи иммуноглобулина (рис. 33.24). Где же находится ДНК, которая кодирует последние 13 остатков вариабельной области? В эмбриональных клетках этот отрезок ДНК локализован в неожиданном месте: вблизи гена С. Указанный отрезок ДНК назвали геном J (от англ. join - соединять), потому что в дифференцированных клетках он соединяет гены V и С. В сущности, в эмбриональных клетках вблизи гена С локализована целая группа расположенных тандемом генов J (рис. 33.25). При дифференцировке клеток, продуцирующих антитела, происходит транслокация гена V в участок, расположенный рядом с геном С; эта транслокация осуществляется путем внутрихромосомной рекомбинации. При этом ген V сращивается (сплайсинг) с геном J и формируется общий ген, полностью кодирующий вариабельный участок. Каждый из указанных генов содержит короткую па- линдромную (разд. 24.27) последовательность, расположенную рядом с участком рекомбинации. Эти палиндромы служат, вероятно, элементами узнавания в процессе рекомбинации.
Рис. 33.24. Ген V, выделенный из эмбриональных клеток, укорочен. Он не кодирует последние 13 аминокислотных остатков вариабельной (V) области полипептидной цепи
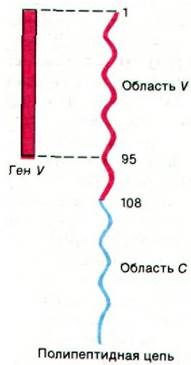
Рис. 33.25. Расположение тандемом группы генов J, кодирующих часть последнего гипервариабельного участка вариабельной области; гены J локализованы вблизи гена С
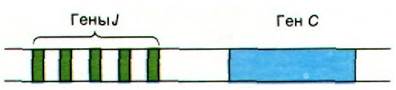
Гены J вносят большой вклад в разнообразие антител, так как они кодируют часть последнего гипервариабельного участка L- и Н-целей. При формировании полного гена Vӽ любой из нескольких сотен генов V может присоединиться к любому из пяти генов J. Например, в результате комбинации 300 неполных генов V с пятью генами J может образоваться 1500 вариантов полного (непрерывного) гена V. Следовательно, соматическая рекомбинация этих генных сегментов усиливает разнообразие, заложенное уже в клетках зародышевого пути.