БІОХІМІЯ - Підручник - Остапченко Л. І. - 2012
Розділ 12. МЕТАБОЛІЗМ ВУГЛЕВОДІВ
12.1.Катаболічні перетворення вуглеводів
12.1.3.Гліколіз
Основними шляхами розщеплення глюкози є гліколіз, пентозо- фосфатний і фосфокетолазний шляхи.
Гліколіз - це послідовність ферментативних реакцій розщеплення глюкози та інших субстратів до пірувату або лактату, що супроводжується біосинтезом АТФ. Цей катаболічний процес відкрито в першій половині минулого століття завдяки працям Г. Ембдена, О. Мейергофа, Я. Парнаса, тому його друга назва - шлях Ембдена - Мейергофа - Парнаса. Гліколіз вважається центральним шляхом катаболізму вуглеводів, що наявний у клітинах тварин, рослин і більшості мікроорганізмів. Залежно від умов існування клітини здатні розщеплювати вуглеводи до пірувату (аеробний гліколіз) або до лактату, пропіонату, бутирату, етанолу, гліцеролу тощо (анаеробний гліколіз).
Анаеробне розщеплення цукрів для отримання метаболічної енергії, яке здійснюється анаеробними організмами, називається бродінням. Залежно від кінцевого продукту процесу можливе молочнокисле, пропіоновокисле, маслянокисле, спиртове та інше бродіння. Анаеробний гліколіз має місце також у деяких тканинах аеробних організмів, зокрема в еритроцитах, сітківці ока, мозковому шарі нирок та скелетному м'язі, що інтенсивно скорочуються, та інших клітинах тварин, які постійно або тимчасово перебувають в умовах гіпоксії.
В аеробних умовах у переважній більшості клітин гліколіз є першим етапом повного окиснення вуглеводів до кінцевих метаболітів - молекул СО2 і Н2О. У самому процесі гліколітичного розщеплення глюкози та інших субстратів, який відбувається в цитоплазмі, можна виділити дві стадії:
✵ активація молекул субстрату (підготовча стадія);
✵ стадія накопичування енергії.
У реакціях першої стадії молекули субстратів активуються шляхом фосфорилювання і перетворюються в один загальний проміжний метаболіт - гліцеральдегід-3-фосфат. На другій стадії молекули гліцеральдегід-3-фосфату окиснюються до пірувату, що супроводжується акумулюванням частини енергії окиснення в молекулах АТФ і НАДН (рис. 12.6).
У ферментативних реакціях першої стадії гліколізу молекули глюкози послідовно фосфорилюються за рахунок АТФ. Спочатку за дії гексокінази в шостому тетраедрі, а після перетворення глюкозо-6-фосфату фосфогексоізомеразою на фруктозо-6-фосфат за участю фосфофруктокінази фосфорилюється в першому тетраедрі молекули з утворенням фруктозо-1,6-дифосфату:
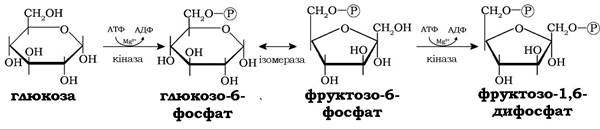
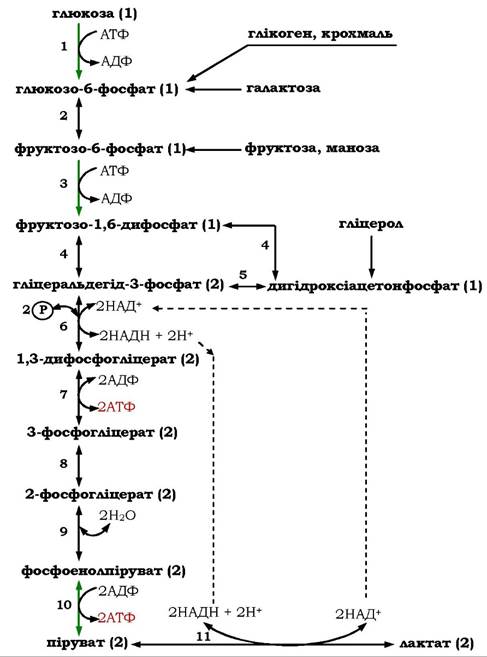
Рис. 12.6. Гліколітичний шлях розщеплення глюкози та інших субстратів гліколізу:
1 - гексокіназа; 2 - фосфогексоізомераза; 3 - фосфофруктокіназа;4 - альдолаза; 5 - тріозофосфатізомераза;
6 - гліцеральдегідфосфатдегідро- геназа; 7 - фосфогліцераткіназа; 8 - фосфогліцеромутаза; 9 - енолаза;
10 - піруваткіназа; 11 - лактатдегідрогеназа.
У дужках праворуч від назв метаболітів гліколізу вказано кількість молекул, які беруть участь у реакції
Обидві реакції фосфорилювання за фізіологічних умов є необоротними. Закінчується підготовча стадія гліколізу розщепленням фруктозо-1,6-дифосфату альдолазою на гліцеральдегід-3-фосфат і дигідроксіацетонфосфат. І хоча у стані динамічної рівноваги цієї оборотної реакції частка дигідроксіацетонфосфату становить 95 %, він швидко перетворюється за дії тріофосфатізомерази на гліцеральдегід-3-фосфат, оскільки подальших перетворень у наступних реакціях гліколізу зазнають лише молекули гліцеральдегід-3-фосфату:
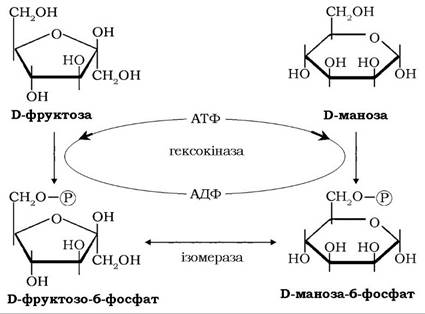
Молекули інших гексозманози, фруктози й галактози також залучаються в гліколіз на цій стадії шляхом їхнього фосфорилювання й подальшого перетворення на гліцеральдегід-3-фосфат.

У клітинах скелетних м'язів, нирок, кишечнику та інших периферійних тканин фруктоза і маноза фосфорилюються за участю гексокінази по вуглецю-6 з утворенням фруктозо-6-фосфату й манозо-6-фосфату, який перетворюється фосфоманозоізомеразою також на фруктозо-6-фосфат:
D-Галактоза залучається до гліколізу довшим за інші гексози шляхом. Спочатку галактокіназа фосфорилює її в галактозо-1- фосфат, який далі в обмінній реакції з уридиндифосфатглюкозою (УДФГ), що каталізується галактозофосфатуридилтрансферазою утворює глюкозо-1-фосфат і уридиндифосфатгалактозу (УДФГал). У наступній реакції залишок галактози в УДФГал перебудовується УДФ-глюкозо-4-епімеразою на глюкозильний залишок УДФГ, яка може взаємодіяти з іншою молекулою галактозо- 1-фосфату або розщеплюватися під впливом УДФ-глюкозопірофосфорилази на глюкозо-1-фосфат і УТФ. Молекули глюкозо-1- фосфату, утворенні в цих реакціях, перетворюються за участю фосфоглюкомутази на глюкозо-6-фосфат:

Крім гексоз субстратом гліколізу може бути і гліцерол - один із продуктів ферментативного гідролізу триацилгліцеролів ендогенного та екзогенного походження. Перетворення гліцеролу в проміжний метаболіт гліколізу - дигідроксіацетонфосфат здійснюється головним чином у печінці за участю двох ферментів: гліцеролкінази і гліцерофосфатдегідрогенази, які послідовно фосфорилюють і окиснюють гліцерол:

У печінці хребетних фруктоза фосфорилюється фруктокіназою по вуглецю-1. Утворений фруктозо-1-фосфат далі розщеплюється альдолазою на дигідроксіацетонфосфат і гліцеральдегід, який за дії гліцеральдегідфосфокінази фосфорилюється в первинній спиртовій групі з утворенням гліцеральдегід-3-фосфату:

Отже, молекули субстратів гліколізу в реакціях підготовчої стадії трансформуються в єдиний метаболіт - гліцеральдегід-3-фосфат і супроводжуються ці зміни витрачанням двох молекул АТФ на активацію молекули гексози або однієї фосфатної групи АТФ на фосфорилювання молекули гліцеролу.
У реакціях другої стадії гліколізу альдегідна група гліцеральдегід-3-фосфату окиснюється НАД-залежною гліцеральдегідфосфатдегідрогеназою. Енергія окиснення, яка спочатку акумулюється в макроергічному фосфоефірному зв'язку при атомі вуглецю-1 молекули 1,3-дифосфосфогліцерату, з високоенергетичною фосфатною групою переноситься фосфогліцераткіназою на молекулу АДФ з утворенням першої молекули АТФ і 3-фосфогліцерату:

Обидві ці спряжені оборотні реакції гліколізу називаються реакцією субстратного фосфорилювання (або фосфорилювання на рівні субстрату), коли фосфорилювання молекули АДФ здійснюється за рахунок енергії, котра вивільняється при окисненні певного субстрату (гліцеральдегід-3-фосфату).
У наступних оборотних реакціях фосфатна група в молекулі 3-фосфогліцерату за допомогою фосфогліцеромутази мігрує до вуглецю-2 з утворенням 2-фосфогліцерату. Подальша дегідратація 2-фосфогліцерату за дії енолази приводить до внутрішньомолекулярного перерозподілу енергії з утворенням макроергічного фосфоефірного зв'язку молекули фосфоенолпірувату:

Перенесення піруваткіназою цієї високоенергетичної фосфатної групи від фосфоенолпірувату на АДФ зумовлює утворення другої молекули АТФ і енольної форми пірувату, яка спонтанно трансформується в кетоформу, котра домінує за фізіологічних значень рН:

Значна втрата стандартної вільної енергії молекули субстрату в піруваткіназній реакції (ΔΟ°'= -31,4 кДж/моль) забезпечує субстратне фосфорилювання молекули АДФ і необоротність цієї реакції. Сумарне рівняння перетворення глюкози в піруват
![]()
після деяких скорочень в обох частинах виглядає наступним чином:
![]()
Отже, у разі гліколітичного перетворення молекули глюкози на дві молекули пірувату утворюються дві молекули АТФ і дві молекули НАДН. НАДН, що генерується у гліцеральдегідфосфатдегідрогеназній реакції, в аеробних умовах переносить відновні еквіваленти на електронтранспортувальну систему мітохондрій, а в анаеробному середовищі (або при гіпоксії) бере участь у відновленні пірувату.
Подальша доля пірувату - одного з головних проміжних метаболітів - залежить у першу чергу від умов існування клітин та їхнього енергетичного потенціалу.
У деяких клітинах тварин (наприклад, у клітинах скелетних м'язів, що інтенсивно скорочуються) в умовах розвитку значної гіпоксії, а також у молочнокислих бактерій піруват відновлюється за дії лактатдегідрогенази до лактату:
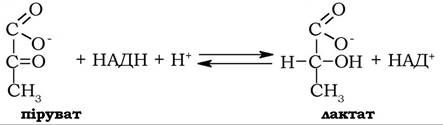
Молекули НАД+, що регенеруються в цій реакції, залучаються до гліцеральдегідфосфатдегідрогеназної реакції. Після відновлення в ній до НАДН, вони реокиснюються в лактатдегідрогеназній реакції (рис. 12.6).
Існування гліколітичного оксидоредуктазного циклу зумовлює можливість перебігу гліколізу в анаеробних умовах, де органічні сполуки діють як донори і як акцептори гідрид-іонів (Н-). Тому в сумарному рівнянні анаеробного гліколізу немає ознак окисно- відновних етапів:
Таким чином, анаеробне розщеплення молекули глюкози на дві молекули лактату супроводжується біосинтезом (у реакціях субстратного фосфорилювання) чотирьох молекул АТФ. За винятком двох молекул АТФ, витрачених на фосфорилювання глюкози і фруктозо-6-фосфату, добуток становить тільки дві молекули АТФ.
У стандартних умовах в ендергонічній реакції
![]()
зміна стандартної вільної енергії ΔG0' = 61,2 кДж/моль; в екзергонічному процесі перетворення![]() глюкози на молочну кислоту
глюкози на молочну кислоту
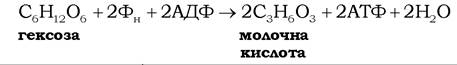
ΔΘ0' дорівнює -196,9 кДж/моль. Отже ефективність гліколізу за стандартних умов становить [(61,2/196,9)· 100%] ~ 31%. Але в клітинах за фізіологічних концентрацій глюкози, лактату, АТФ, АДФ і неорганічного фосфату ефективність акумулювання у фосфоефірних зв'язках молекули АТФ звільненої в ході гліколізу вільної енергії, є вдвічі більшою (>60 %).
Утворення лактату за нормальних умов життєдіяльності спостерігається також у клітинах шлунково-кишкового тракту, мозку та в еритроцитах. Енергетика останніх за відсутності мітохондрій практично забезпечується гліколізом навіть в аеробних умовах.
У клітинах різних тканин лактатдегідрогеназа (ЛДГ), яка є олігомерним білком з молекулярною масою 134 кДа, представлена п'ятьма ізоферментними формами - тетрамерами, утвореними різними комбінаціями двох поліпептидних субодиниць Н і М. Так, у клітинах міокарда фермент представлений ізоформою ЛДГ1, складеною чотирма Н-доменами (Н4), яка виявляє більшу спорідненість до лактату, сприяючи його повному окисненню в мітохондріях серцевого м'яза. Натомість ізофермент ЛДГ5, локалізований у клітинах скелетних м'язів, є тетрамером (М4), що вмішує лише М-поліпептидні ланцюги. Цей ізофермент має більшу спорідненість до пірувату і швидко відновлює його до лактату, котрий з кров'ю потрапляє до печінки, де реокиснюється в піруват під дією ізоферменту ЛДГ3 печінки, що вмішує одну Н- і три М-субодиниці (НМ3). Отже, домінуючий напрямок лактатдегідрогеназної реакції залежить від локалізації в клітинах різних ізоформ ферменту з різною субстратною специфічністю. При пошкодженні тканин вміст ферменту в крові зростає і це дає можливість визначати наявність і ступінь ураження органів, зокрема міокарда, за оцінкою ізоферментного спектра лактатдегідрогенази.
Бродіння. У клітинах деяких мікроорганізмів (зокрема, дріжджів) піруват, що утворюється в процесі анаеробного гліколізу, за відсутності ферменту лактатдегідрогенази відновлюється в етанол. У цих клітинах замість лактатдегідрогеназної реакції проходять дві спряжені ферментативні реакції, в яких піруват спочатку за участю піруватдекарбоксилази необоротно перетворюється в ацетальдегід, а останній відновлюється алкогольдегідрогеназою до етанолу:
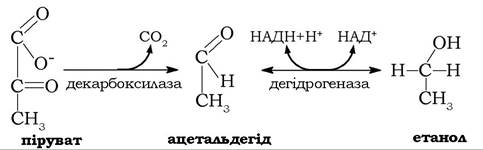
Донором відновних еквівалентів в другій реакції слугує НАДН, що був генерований у гліцеральдегідфосфатдегідрогеназній реакції гліколізу (рис. 12.6).
Продукти спиртового бродіння етанол і СО2 є кінцевими продуктами катаболізму цукрів у цих мікроорганізмів і тому виділяються в навколишнє середовище. Сумарне рівняння спиртового бродіння має такий вигляд:
![]()
Таким чином, ферментативне розщеплення глюкози та інших субстратів в анаеробних умовах до лактату або етанолу, хоча й включає окисно-відновні реакції, але зміни ступеня окиснення вуглецю в сумарних реакціях молочнокислого та спиртового бродіння немає. Іншими словами, перетворення молекул глюкози в молекули лактату або етанолу та СО2, що супроводжується звільненням і акумулюванням частини вільної енергії молекули глюкози у фосфатних групах молекул АТФ, здійснюється без окиснення атомів вуглецю в молекулах кінцевих продуктів анаеробного гліколізу. Незважаючи на невеликий енергетичний вихід (~ 7 % від загальної вільної енергії молекули глюкози), анаеробний гліколіз відносять до унікального за ефективністю процесу серед відомих катаболічних перетворень речовин, заснованих майже виключно на окисненні органічного субстрату, оскільки він дозволяє клітинам синтезувати АТФ в анаеробних умовах.
Регуляція гліколізу. Швидкість перебігу гліколізу визначається насамперед енергетичним потенціалом клітин, а також доступністю субстратів. Включення глюкози, інших гексоз і глікогену в гліколіз (глікогеноліз) регулюється в основному активністю гексокінази та глікогенфосфорилази, котрі каталізують перші етапи гліколізу та глікогенолізу відповідно. Як відомо, активність цих ферментів контролюється механізмами алостеричної модуляції та ковалентної модифікації: гексокіназа скелетних м'язів алостерично гальмується власним продуктом - глюкозо-6-фосфатом і АТФ, а активність глікогенфосфорилази може контролюватися і алостеричним модуляторами (АТФ і АМФ), і шляхом фосфорилювання-дефосфорилювання залишків серину в активних центрах ферменту.
Як правило, необоротні реакції метаболічних шляхів каталізуються регуляторними ферментами. У гліколізі такими є три кіназні реакції. Крім гексокінази в регуляції швидкості гліколізу беруть участь також фосфофруктокіназа й піруваткіназа, алостеричними модуляторами для якої слугують АТФ, ацетил-КоА, жирні кислоти, фруктозо-1,6-дифосфат.
Фосфофруктокіназа, яка вважається ключовим регуляторним ферментом гліколізу в скелетних м'язах, відрізняється від інших кіназ цього шляху складністю будови та регуляції активності. Фермент має декілька алостеричних центрів, розташованих на чотирьох субодиницях. Його алостеричними інгібіторами є АТФ, цитрат, жирні кислоти. Тому в стані спокою у м'язах при високому енергетичному потенціалі (високому співвідношенні АТФ/АМФ) і високої концентрації цитрату й жирних кислот, активність ферменту є мінімальною. Інгібуючий вплив АТФ, цитрату й жирних кислот знімається з підвищенням концентрації продуктів гідролізу АТФ - АДФ і, особливо, АМФ, що має місце під час активного скорочення скелетних м'язів.
В інших органах і тканинах, зокрема в печінці, мозку, нирках тощо, головним алостеричним регулятором гліколізу є фруктозо- 2,6-дифосфат, який одночасно може активувати фосфофруктокіназу і гальмувати активність фруктозо-1,б-дифосфатази - регуляторного ферменту глюконеогенезу.
Концентрація фруктозо-2,6-дифосфату в клітинах контролюється біфункціональним ферментом б-фосфофрукто-2-кіназо/фруктозо-2,б-дифосфатазою, що каталізує реакції синтезу й дефосфорилювання фруктозо-2,6-дифосфату:
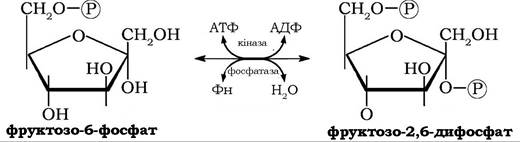
Активуючий ефект фруктозо-2,6-дифосфату на фосфофруктокіназу знімається дефосфорилюванням фруктозо-2,6-дифос- фату. Важливість регуляторної ролі фруктозо-2,6-дифосфату в гліколізі підтверджується тим фактом, що регуляція внутрішньоклітинного вмісту цього метаболіту здійснюється на рівні індукції альтернативних ізоформ ферменту з кіназною або фосфатазною активністю.
Отже, швидкість гліколізу контролюється його регуляторними ферментами-кіназами, що каталізують лімітуючі етапи процесу. Активність гексокінази, фосфофруктокінази й піруваткінази модулюється алостерично: активаторами (АМФ, АДФ, фруктозо- 1,6- і -2,6,-дифосфатами) та інгібіторами (глюкозо-6-фосфатом, АТФ, цитратом, жирними кислотами, ацетил-КоА та ін.).
Домінування того чи іншого типу модуляторів залежить головним чином від енергетичного потенціалу клітин (АТФ/АМФ), а також від забезпеченості проміжними метаболітами (цитрат, ацетил-КоА, глюкозо-6-фосфат тощо) для використання в біосинтетичних реакціях.
Необхідно відмітити, що в умовах оксигенації тканин спостерігається глибоке гальмування анаеробного гліколізу, яке називається ефектом Пастера. Це явище пов'язано з активацією клітинного дихання, тобто процесом повного окиснення пірувату в циклі трикарбонових кислот.
В еритроцитах ссавців разом із гліколітичним перетворенням глюкози має місце дифосфогліцератний цикл (рис. 12.7), в якому утворюється 2,3-дифосфогліцерат, котрий відіграє важливу роль у регуляції транспорту кисню гемоглобіном. Ця сполука може синтезуватися двома шляхами (шунт Рапопорта): ізомеризацією 1,3-дифосфогліцерату за участю дифосфогліцератмутази та фосфорилюванням 3-фосфогліцерату за дії фосфогліцераткінази. На другому шляху проходить і оборотна реакція, яка каталізується фосфогліцератфосфатазою.
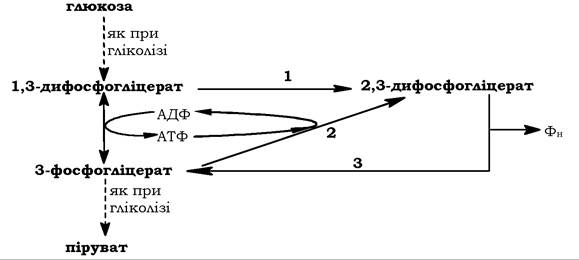
Рис. 12.7. Утворення 2,3-дифосфогліцерату при перетворенні глюкози:
1 - дифосфогліцератмутаза; 2 - фосфогліцераткіназа; 3 — фосфогліцератфосфатаза
Присутність 2,3-дифосфогліцерату в еритроцитах знижує спорідненість гемоглобіну до кисню. Це сприяє дисоціації кисню з оксигемоглобіну й поглинання його тканинами.
Окисне декарбоксилювання пірувату. Основним енергозначимим перетворенням пірувату в аеробних умовах є його окисне декарбоксилювання з утворенням ацетил-КоА - вихідної сполуки циклу трикарбонових кислот (ЦТК):
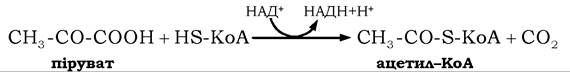
Цей процес здійснюється в клітинах еукаріотів у мітохондріях, куди піруват потрапляє з цитоплазми за допомогою специфічного переносника.
Одночасне декарбоксилювання і дегідрування піруваїу каталізується мультиензимним піруватдегідрогеназним комплексом, локалізованим у матриксі мітохондрій рослин і тварин або прикріпленим до клітинної стінки у мікроорганізмів. Центральним у комплексі, який складається із трьох ферментів і п'яти коферментів, є фермент дигідролшоатацетилтрансфераза, з яким асоційовані молекули піруватдегідрогенази і дигідроліпоатдегідрогенази.
На першому етапі (1) процесу піруватдегідрогеназа [Е1] каталізує відщеплення карбоксилу від молекули пірувату й перенесення гідроксіетильного залишку на кофермент тіамінпірофосфат (ТПФ) з утворенням гідроксіетил-ТПФ (рис. 12.8). Наступне дегідрування гідроксіетил-ТПФ і перенесення атомів водню та ацетилу на простетичні групи центрального ферменту комплексу (2) також відбувається за участю піруватдегідрогенази.

Рис. 12.8. Етапи окисного декарбоксилювання пірувату, який каталізується ферментами піруватдегідрогеназного комплексу
Ліпоатамідні простетичні групи дигідроліпоатацетилтрансферази [Е2], які утворюються в результаті приєднання карбоксильних груп молекул ліпоєвої кислоти до аміногруп специфічних залишків лізину в активних центрах ферменту, виконують у цьому комплексі роль переносників ацетильних груп і атомів водню між простетичними групами двох інших ферментів комплексу (рис. 12.8).
Приєднання атомів водню та ацетильної групи до окисненої (дисульфідної) форми ліпоатамідних груп дигідроліпоатацетилтрансферази супроводжується утворенням відновленої ацетильованої форми ліпоатамідних груп ферменту. Далі ацетильна похідна дигідроліпоатацетилтрансферази взаємодіє з вільним КоА-SH (3) з утворенням молекули ацетил-КоА і повністю відновленої (дитіоло- вої) форми простетичних груп. На наступному етапі (4) дитіолові ліпоатамідні групи ферменту [Е2] окиснюються дигідроліпоатдегід- рогеназою [Е3] з утворенням вихідної дисульфідної форми простетичних груп дигідроліпоатацетилтрансферази. Атоми водню, котрі відщеплюються дигідроліпоатдегідрогеназою, спочатку переносяться на її кофермент ФАД, після чого, на завершальному етапі (5), передаються від ФАДН2 на молекулу НАД+. Відновлений НАДН спрямовує атоми водню на дихальний ланцюг (ДЛМ), розташований на внутрішній мембрані мітохондрій. Молекула ацетил-КоА, що утворилася з пірувату в реакціях піруватдегідрогеназного комплексу, може або окиснюватися в циклі трикарбонових кислот у матриксі мітохондрій, або транслокуватися (у формі цитрату) у цитоплазму для участі в біосинтезі вищих жирних кислот, холестеролу, кетонових тіл тощо.
Процес окисного декарбоксилювання пірувату, що є необоротним у клітинах тварин, регулюється механізмами ковалентної модифікації та алостеричної модуляції. За високої концентрації у тканинах АТФ, ацетил-КоА і проміжних продуктів ЦТК активується один із двох регуляторних ферментів піруватдегідрогеназного комплексу - кіназа піруватдегідрогенази, яка інгібує активність піру- ватдегідрогенази шляхом фосфорилювання залишків серину в її активному центрі. Інгібуючий ефект фосфорилювання серину знімається його дефосфорилюванням за дії другого регуляторного ферменту комплексу - фосфатази фосфопіруватдегідрогенази, що стимулюється підвищенням концентрації іонів Са2+, яке зазвичай сигналізує про зниження відношення АТФ/ АМФ у клітинах.
Швидкість окисного декарбоксилювання пірувату регулюється також механізмом алостеричної модуляції. Молекули ацетил-КоА і НАДН, котрі утворюються в мітохондріях при окисненні пірувату й вищих жирних кислот, є алостеричними інгібіторами піруват- дегідрогеназної реакції. Тому в абсорбтивний період або під час іммобілізації жирних кислот із жирових депо клітин спостерігається гальмування процесу перетворення пірувату в ацетил-КоА.