БІОХІМІЯ - Підручник - Остапченко Л. І. - 2012
Розділ 16. ЕНЕРГЕТИЧНИЙ ОБМІН
16.4. Цикл трикарбонових кислот
Цикл трикарбонових кислот (ЦТК, цикл цитринової кислоти або цикл Кребса) - загальний шлях окиснення вуглеводів, білків і ліпідів, оскільки в ході метаболізму глюкози, амінокислот і жирних кислот утворюється ацетил-КоА (рис. 16.17). У цьому циклі, який в еукаріотичних клітинах відбувається в мітохондріях, ацетат окиснюється до СО2 і саме О2є кінцевим акцептором електронів. Також бере участь ЦТК у процесах глюкогенезу, переамінування, дезамінування й ліпогенезу.
Починається ЦТК із взаємодії ацетил-КоА та оксалоацетату за дії ферменту цитратсинтетази, що приводить до утворення цитрату. Ця реакція є трифазною. Спочатку ацетил-КоА перетворюється в енольну форму:

Потім ця форма ацетил-КоА вступає в реакцію з оксалоацетатом і утворюється цитрил-КоА:

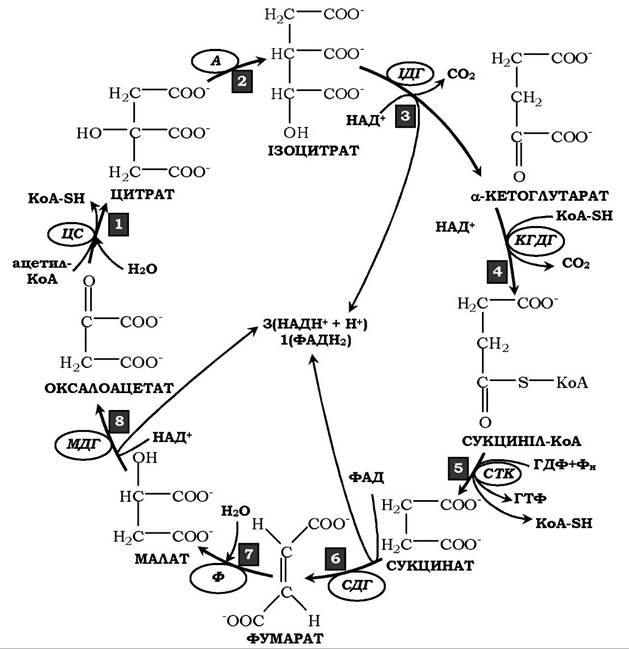
Рис. 16.17. Загальна схема циклу трикарбонових кислот:
1 - утворення цитрату; ЦС - цитратсинтаза; 2 - перетворення цитрату в ізоцитрат; А - аконітаза; 3 - окисне декарбоксилювання ізоцитрату; ІДГ - ізоцитратдегідрогеназа; 4 - окисне декарбоксилювання а-кетоглугарату; α-КГДГ - а-кетоглутаратдегідрогеназа; 5 - перетворення сукциніл-КоА в сукцинат; СТК - сукцинаттіокіназа; 6 - дегідрування сукцинату; СДГ - сукцинатдегідрогеназа; 7 - утворення малату з фумарату;
Ф - фумараза; 8 - дегідрування малату; МДГ — малатдегідрогеназа
Далі відбувається утворення цитрату при гідролізі тіоефірного зв'язку й вивільнення коферменту А за участю ферменту цитрат- синтетази:
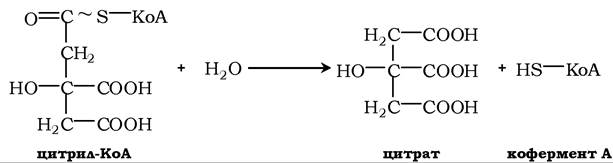
Ця реакція характеризується відносно великими витратами енергії у вигляді тепла, що зумовлює зміщення її рівноваги в бік утворення цитрату.
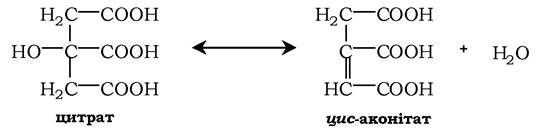
На подальшій стадії цитрат перетворюється в цис-аконітат у реакції дегідратації. Ця реакція каталізується ферментом аконітатгі- дролазою (аконітазою), яка містить ферум у Ре2+-стані:
Наступне утворення ізоцитрату теж каталізується аконітатгідролазою в реакції гідратації:
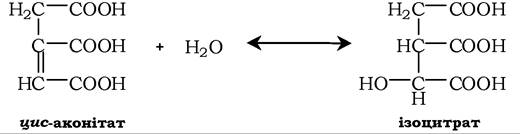
Існує думка, що цис-аконітат може бути не обов'язково інтермедіатом між цитратом та ізоцитратом, а утворюється на боковій гілці основного шляху.
Фермент ізоцитратдегідрогеназа каталізує реакцію дегідрування, в якій утворюється з ізоцитрату оксалосукцинат. Цей самий фермент каталізує утворення з останнього α-кетоглутараіу в реакції декарбоксилювання за участю іонів Мn2+ (або Mg2+). Відомі три форми цього ферменту. Одна з них НАД+-залежна, знайдена тільки в мітохондріях і відіграє провідну роль в окисненні ізоцитрату. Дві інші НАДФ+-залежні, одна з яких локалізована в мітохондріях, а інша в цитозолі. Вони, мабуть, відіграють допоміжну роль.
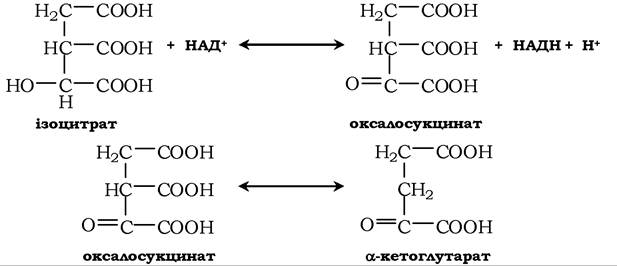
За дії α-кетоглутаратдегідрогеназного комплексу (потребує участі тіамінопірофосфату, ліпоату, НАД+, ФДД і КоА) α-кетглутарат при окисному декарбоксилюванні перетворюється в сукциніл-КоД з високоенергетичним зв'язком. За фізіологічних умов ця реакція необоротна. Механізм цієї реакції подібний до окисного декарбоксилювання пірувату.
У подальшому фермент сукцинаттіокіназа за участю Mg2+ каталізує перетворення сукциніл-КоД в сукцинат і ця реакція спряжена з утворенням ГТФ при фосфорилюванні ГДФ. Це єдина стадія циклу трикарбонових кислот, в якій утворюється високоенергетичний зв'язок на так званому субстратному рівні.
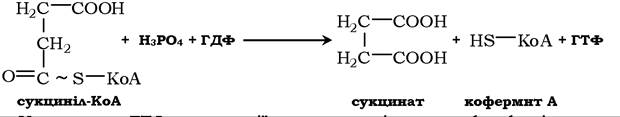
Утворення ГТФ у реакції, яка каталізується фосфокіназою, може приводити до синтезу ДТФ:

Фермент сукцинатдегідрогеназа, який містить ФАД і залізосірчаний білок (Fc-S) каталізує перетворення сукцинату при дегідруванні у фумарат:

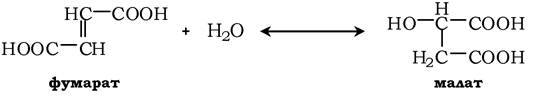
Перетворення фумарату в малат у реакції гідратації каталізує фермент фумаратгідратаза (фумараза):
![]()
Утворений малат в НДД+-залежній реакції, яка каталізується ферментом малатдегідрогеназою, перетворюється в оксалоацетат і, таким чином, один оберт ЦТК завершується:

Слід відмітити, що реакції ЦТК є оборотними, але при перетворенні цитрил-КоД в цитарт і α-кетоглутарату в сукциніл-КоД рівновага значно зсунута в бік утворення останніх. Саме тому ці реакції за фізіологічних умов можна вважати однонаправленими.
Під час кожного повторення ЦТК атоми вуглецю надходять до нього у вигляді ацетил-КоД, а виводяться у вигляді СО2. В окисно-відновних реакціях утворюється чотири пари атомів водню (з яких три акцептовані у вигляді НДДН+ Н+, а одна - ФДДН2), що надходять у дихальний ланцюг мітохондрій.
Слід зауважити, що ферменти ЦТК за винятком α-кетоглутаратдегідрогенази та сукцинатдегідрогенази, містяться також і поза мітохондріями. Але деякі з цих ферментів, зокрема малатдегідрогеназа, мають відмінності від відповідних ферментів мітохондрії.
У рослин і мікроорганізмів поряд зі звичайним ЦТК, функціонує його модифікуючий варіант - гліоксилатний цикл. Перетворення ізоцитрату в цьому циклі каталізує ізоцитратліаза, продукти цієї реакції гліоксилат і сукцинат. У подальшому гліоксилат взаємодіє з ацил-КоД й утворена сполука перетворюється в малат. У свою чергу, сукцинат через фумарат і малат перетворюється в оксалоацетат. У мікроорганізмів існують ще інші модифікації циклу трикарбонових кислот.
У циклі трикарбонових кислот за один його оберт на кожну перетворену молекулу ацетил-КоА утворюються три молекули НАДН і одна молекула ФАДН2. У процесі окисного фосфорилювання, який відбувається за участю внутрішньої мембрани мітохондрій, на кожну молекулу НАДН синтезується три молекули АТФ з АДФ і Фн, а ФАДН2 - дві молекули АТФ. Таким чином, 3 молекули НАДН і 1 молекула ФАДН2 сприяють утворенню 11 молекул АТФ. Крім того, з ГТФ, який утворився з ГДФ і Фн при перетворені сукциніл-КоА в сукцинат, у реакції, що каталізується фосфокіназою, утворюється АТФ.
Підсумок кількості утворених молекул АТФ за участю ЦТК і окисного фосфорилювання свідчить про можливість синтезу 12 молекул АТФ.
Унаслідок повного окиснення однієї молекули ацетил-КоА вивільняється енергії близько 898,7 кДж/моль. Разом із тим, на синтез 12 молекул АТФ витрачається енергії 367,2 кДж/моль. Отже, коефіцієнт корисної дії (η) ЦТК становить приблизно 40 %:
![]()
Крім енергетичної функції ЦТК відіграє роль постачальника метаболітів для синтезу ряду речовин. Зокрема, α-кетоглутарат може бути попередником проліну, глутаміну, глутамату; сукциніл-КоА - попередником гему в гемоглобіні; оксалоацетат - попередник як вуглеводів, так і аспарагіну й аспартату.