Учебник Биология - ВУНМЦ 2000
Глава 4. ОРГАНИЗМЕННЫЙ УРОВЕНЬ ОРГАНИЗАЦИИ ЖИВОГО
4.2. БИОЛОГИЯ РАЗВИТИЯ
4.2.1. ОНТОГЕНЕЗ (ИНДИВИДУАЛЬНОЕ РАЗВИТИЕ ОСОБИ)
Онтогенез - индивидуальное развитие особи - начинается с момента слияния сперматозоида с яйцеклеткой и образования зиготы, заканчивается смертью.
Есть два типа онтогенеза: прямой и непрямой. Непрямое развитие может быть личиночным, прямое развитие - в двух формах: неличиночной и внутриутробной.
Личиночная форма характеризуется наличием одной или нескольких личиночных стадий. Личинки активно питаются, имеют временные органы. Этот тип развития сопровождается метаморфозом.
Неличиночная форма развития характерна для рыб, рептилий, птиц. Яйца этих животных богаты желтком. Для дыхания, выделения, питания развивающегося зародыша есть временные органы.
Внутриутробная форма характерна для млекопитающих и человека. Все функции зародыша осуществляются за счет организма матери, с помощью специального органа - плаценты.
4.2.1.1. Эмбриональное развитие
При любой форме онтогенеза выделяют следующие периоды: эмбриональный и постэмбриональный. До эмбрионального периода происходит гаметогенез - образование сперматозоидов и яйцеклеток. Эмбриональный период имеет следующие этапы:
1) Дробление - образование бластулы;
2) Гаструляция - образование зародышевых листков;
3) Гисто- и органогенез - образование органов и тканей зародыша.
При личиночной форме онтогенеза эмбриональный период начинается с образования зиготы и заканчивается выходом из яйцевых оболочек.
При неличиночной форме онтогенеза эмбриональный период начинается с образования зиготы и заканчивается выходом из зародышевых оболочек.
При внутриутробной форме онтогенеза эмбриональный период начинается с образования зиготы и длится до рождения.
4.2.1.1.1. Дробление
В результате оплодотворения образуется зигота, которая начинает дробиться. Дробление сопровождается митотическим делением. Нет роста клеток, и объем зародыша не изменяется. Это происходит потому, что между делениями в короткой интерфазе отсутствует постмитотический период, а синтез ДНК начинается в телофазе предшествующего митотического деления. Клетки, образующиеся в процессе дробления, называются бластомерами, а зародыш - бластулой.
Типы дробления зависят от количества и распределения желтка в яйцеклетках (рис. 96).
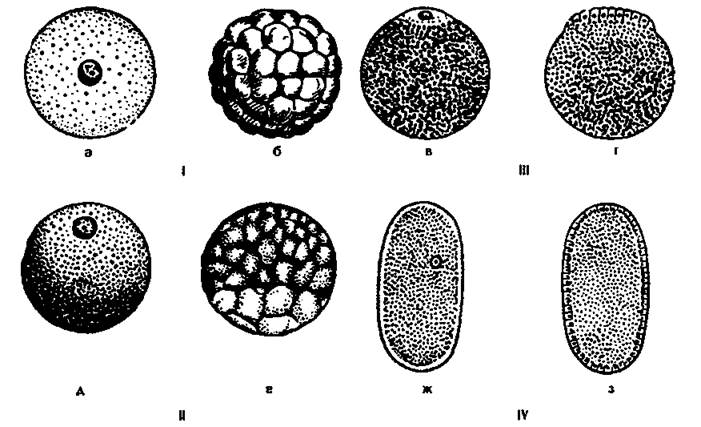
Рис. 96. Типы яиц и характер их дробления (яйца и зародыши изображены вверх анимальным полюсом).
I - изолецитальное яйцо (а) и его равномерное дробление (б), II и III - телолецитальные яйца (в и д) и их дробления - дискоидальное (г) и неравномерное (е), IV - центролецитальное яйцо (ж) и его поверхностное дробление (з).
Дробление может быть:
✵ полным равномерным;
✵ полным неравномерным;
✵ неполным дискоидальным;
✵ неполным поверхностным.
Полное равномерное дробление характерно для изолецитальных яиц, например, для ланцетника. Ядро зиготы делится митозом на два, затем делится цитоплазма. Борозда дробления проходит по меридиану, образуя два бластомера. Затем снова делится ядро, и на поверхности зародыша появляется вторая борозда дробления, идущая по меридиану перпендикулярно первой. Образуются четыре бластомера. Третья борозда проходит по экватору и делит его на восемь частей. Затем происходит чередование меридионального и экваториального дроблений. Число бластомеров увеличивается. Зародыш на стадии 32 бластомеров называют морулой. Дробление продолжается до образования зародыша, похожего на пузырек, стенки которого образованы одним слоем клеток, называемом бластодермой. Бластомеры расходятся от центра зародыша, образуя полость, которая называется первичной или бластоцелью. Бластомеры имеют одинаковые размеры. В результате такого дробления образуется целобластула (рис. 97).
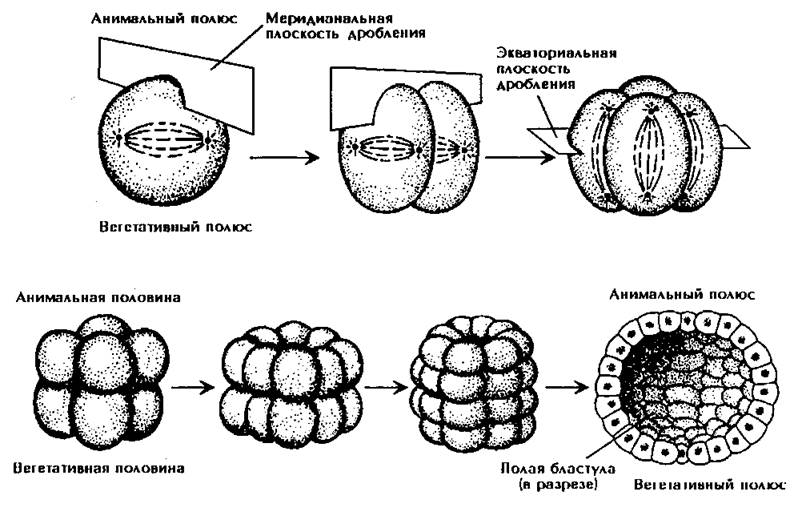
Рис. 97. Голобластическое дробление иглокожего Synapta digits, приводящее к образованию полой бластулы, которая изображена в разрезе во втором ряду справа (по Saunders, 1982).
Полное неравномерное дробление характерно для телолецитальных яиц с умеренным содержанием желтка, например, у лягушки. Первая и вторая борозды дробления проходят по меридианам и полностью делят яйцо на четыре части. Третья борозда смещена в сторону анимального полюса, где нет желтка. Бластомеры имеют неодинаковую величину: на анимальном полюсе они меньше (микромеры), на вегетативном больше (макромеры). Желток затрудняет дробление, и поэтому дробление макромеров идет медленнее, чем микромеров. Стенка бластулы состоит из нескольких рядов клеток. Первичная полость мала и смещена к анимальному полюсу. Образуется амфибластула.
Неполное дискоидальное дробление характерно для телолецитальных яиц с большим содержанием желтка, например, для рептилий, птиц. Дробление идет только на анимальном полюсе. Первая и вторая борозды дробления проходят по меридиану перпендикулярно друг другу. Третья борозда смещена к анимальному полюсу, в результате чего образуется зародышевый диск. Бластоцель располагается под слоем бластодермы в виде щели. Бластула называется дискобластулой (рис. 98).
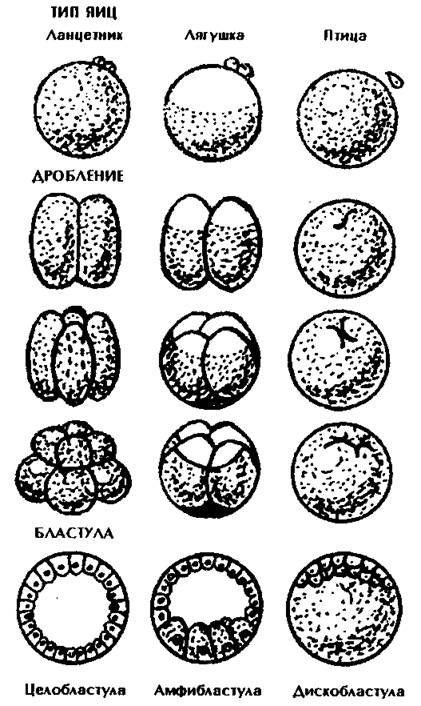
Рис. 98. Типы яиц и соответствующие им типы дробления.
Неполное поверхностное дробление характерно для центролецитальных яиц, например, для членистоногих. Ядра центролецитальных яиц многократно делятся и перемещаются к периферии, где в цитоплазме нет желтка. Образуются бластомеры. Бластула имеет один слой бластомеров. Бластоцель заполнена желтком. Такая бластула называется перибластулой.
4.2.1.1.2. Гаструляция
По окончании периода дробления у многоклеточных животных начинается период образования зародышевых листков - гаструляция. Гаструляция связана с перемещением эмбрионального материала. Сначала образуется ранняя гаструла, имеющая два зародышевых листка (эктодерму и энтодерму), затем поздняя гаструла, когда формируется третий зародышевый листок - мезодерма. Образующийся зародыш называют гаструлой (рис. 99).

Рис. 99. Типы гаструл: а - инвагинационная гаструла Phoronis, б и б' - две стадии развития иммиграционной гаструлы медузы Octorchis gegenbauri, в и в' - две стадии развития деляминационной гаструлы Geryonid, г и г' - две стадии развития эпиболической гаструлы Bonellia. 1 - эктодерма, 2 - энтодерма, 3 - бластоцель.
Образование ранней гаструлы происходит следующим образом:
✵ иммиграцией (выселением клеток), как у кишечнополостных;
✵ инвагинацией (впячиванием), как у ланцетника;
✵ эпиболией (обрастанием), как у лягушки;
✵ деляминацией (расщеплением), как у некоторых кишечнополостных.
При иммиграции (выселении) часть клеток бластодермы с поверхности зародыша уходит в бластоцель. Образуется наружный слой - эктодерма и внутренний - энтодерма. Бластоцель заполнена клетками. Такой способ образования гаструлы характерен для кишечнополостных.
Для ланцетника характерно образование гаструлы путем инвагинации (впячивания). При инвагинации определенный участок бластодермы (вегетативный полюс) прогибается внутрь и достигает анимального полюса. Образуется двухслойный зародыш - гаструла. Наружный слой клеток называют эктодермой, внутренний - энтодермой. Энтодерма выстилает полость первичной кишки (гастроцель). Отверстие, при помощи которого полость сообщается с внешней средой, называется первичным ртом - бластопором. У первичноротых животных (черви, моллюски, членистоногие) он превращается в ротовое отверстие. У вторичноротых - в анальное отверстие, а рот образуется на противоположном конце тела (хордовые).
Эпиболия (обрастание) характерна для животных, развивающихся из телолецитальных яиц. Образование гаструлы идет за счет быстрого деления микромеров, которые обрастают вегетативный полюс. Макромеры оказываются внутри зародыша. Образование бластопора не происходит и нет гастроцели. Эпиболия характерна для амфибий.
Деляминация (расслоение) встречается у кишечнополостных, бластула которых похожа на морулу. Клетки бластодермы делятся на наружный и внутренний слои. Наружный слой образует эктодерму, внутренний - энтодерму.
У всех многоклеточных, кроме губок и кишечнополостных, образуется третий зародышевой листок - мезодерма. Формирование мезодермы происходит двумя способами (рис. 100):
1) Телобластическим;
2) Энтероцельным.
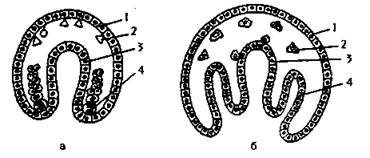
Рис. 100. Схема образования мезодермы (по Шимкевичу, 1925, модифицировано): а - у первичноротых, б - у вторичноротых; 1 - эктодерма, 2 - мезенхима, 3 - энтодерма, 4 - телобласт (а) и целомическая мезодерма (б).
Телобластический способ характерен для первичноротых. На границе между эктодермой и энтодермой по бокам от бластопора клетки - телобласты - начинают делиться и дают начало мезодерме.
Энтероцельный способ характерен для вторичноротых. Клетки, формирущие мезодерму, обособляются в виде карманов первичной кишки. Полости карманов превращаются в целом. Мезодерма делится на отдельные участки - сомиты, из которых образуются определенные ткани и органы.
4.2.1.1.3. Гисто- и органогенез
После образования мезодермы начинается процесс гисто- и органогенеза. Сначала формируются осевые органы - нервная трубка, хорда, затем все остальные.
У ланцетника из эктодермы на спинной стороне зародыша образуется нервная трубка (рис. 101). Остальная эктодерма формирует кожный эпителий и его производные. Из энто- и мезодермы под нервной трубкой формируется хорда. Под хордой находится кишечная трубка энтодермального происхождения. По бокам от хорды - мезодерма сомитов, которая дифференцирована следующим образом: наружная часть сомита, прилегающая к эктодерме, называется дермотомом. Из нее образуется соединительная ткань кожи. Внутренняя часть - склеротом - дает начало скелету. Между дермотомом и склеротомом находится миотом, дающий начало поперечнополосатой мускулатуре. Под сомитами расположены его ножки (нефрогонотом), из которых формируется мочеполовая система. Целомические мешки образуются симметрично по бокам. Внутри целомических мешков находится вторичная полость тела - целом, - заполненная жидкостью. Стенки целомических мешков, обращенные в сторону кишечника, называют спланхноплеврой, в сторону эктодермы - соматоплеврой. Эти листки участвуют в образовании сердечнососудистой системы, плевры, брюшины, перикарда.
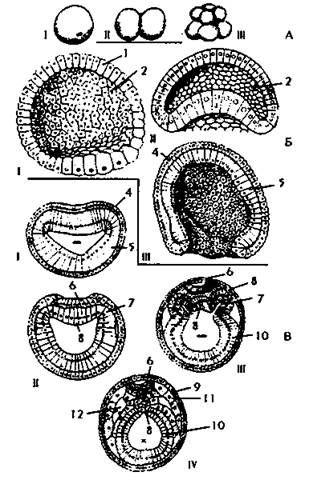
Рис. 101. Эмбриональное развитие ланцетника. А - период дробления: I - зигота, II - двухбластомерная стадия, III - восьмибластомерная стадия. Б - бластула (I), ранняя (II) и более поздняя (III) стадии гаструляции: 1 - бластодерма, 2 - бластоцель (первичная полость тела), 3 - начало впячивания (инвагинации) вегетативной части бластулы, 4 - первичная эктодерма, 5 - первичная энтодерма. В - закладка осевых органов. Поперечные разрезы ранней (I) и более поздней (II) гаструл, ранней (III) и поздней (IV) нейрул: 6 - нервная пластинка, прогибающаяся затем в нервный желобок, 7 - мезодермальные пузыри, 8 - хорда, 9 - кожная эктодерма, 10 - кишечная энтодерма, 11 - первичный сегмент мезодермы, 12 - вторичная полость тела (целомическая). Знаком "-" показана полость первичной кишки (гастроцель), знаком "х" - полость кишечной трубки, в которую замкнулась кишечная энтодерма.
Таким образом, из эктодермы формируются наружный эпителий, кожные железы, эмаль зубов, волосы, ногти, когти.
Из энтодермы образуются эпителий средней кишки, печень, поджелудочная железа, щитовидная железа, тимус, эпителий дыхательной системы.
Мезодерма участвует в образовании мышц, соединительной ткани, костной ткани, каналов выделительной системы, кровеносной системы, части ткани половых желез.