БИОЛОГИЯ Том 1 - руководство по общей биологии - 2004
9. ИСПОЛЬЗОВАНИЕ ЭНЕРГИИ
9.3. Клеточное дыхание
9.3.5. Аэробное дыхание
При аэробном дыхании образующаяся в процессе гликолиза пировиноградная кислота в конечном итоге полностью окисляется кислородом до СО2 и воды. В первой фазе пировиноградная кислота расщепляется с образованием СО2 и водорода. Этот процесс протекает в матриксе митохондрий и включает в себя последовательность реакций, называемую циклом Кребса. Во второй фазе отщепившийся водород через ряд окислительно-восстановительных реакций — в так называемой дыхательной цепи — окисляется в конечном счете молекулярным кислородом до воды. Это происходит на так называемых кристах (гребневидных складках внутренней мембраны митохондрий).
Начальные этапы аэробного дыхания представлены на рис. 9.6.
Переходный этап между гликолизом и циклом Кребса
Каждая молекула пировиноградной кислоты поступает в матрикс митохондрий и здесь — в виде ацетильной группы (СН3СОО—) — соединяется с веществом, которое называется коферментом А (или сокращенно КоА), в результате чего образуется ацетилкофермент А (ацетил-КоА). Ацетильная группа содержит два атома углерода (2С), поэтому для того чтобы она могла образоваться, пировиноградная кислота (3С) должна утратить атом углерода.
Отщепление атома углерода в виде СО2 называется реакцией декарбоксилирования. Это — окислительное декарбоксилирование, поскольку оно сопровождается окислением путем дегидрирования, в результате чего образуется восстановленный НАД.
Цикл Кребса
Этот цикл назван так в честь открывшего его в 1930-х годах исследователя — сэра Ганса Кребса. Его называют также «циклом трикарбоновых кислот» и «циклом лимонной кислоты», поскольку именно эти кислоты в нем участвуют.
Цикл Кребса (рис. 9.6) протекает в матриксе митохондрий. Ацетильные группы (2С) вовлекаются в цикл, присоединяясь к 4С-соединению — щавелево-уксусной кислоте, в результате чего образуется лимонная кислота (6С). Далее следует цикл реакций, в которых поступившие в цикл ацетильные группы декарбоксилируются с образованием двух молекул СО2 и дегидрируются с высвобождением четырех пар атомов водорода, присоединяющихся к переносчикам, в результате чего образуются три молекулы восстановленного НАД и одна молекула восстановленного ФАД. Каждый оборот цикла дает также одну молекулу АТФ. (Напомним, что из одной молекулы глюкозы образуются две ацетильные группы, и значит, для окисления каждой молекулы глюкозы требуются два оборота цикла.) В конце цикла щавелево-уксусная кислота регенерирует и может теперь присоединить к себе новую ацетильную группу.
Общий баланс аэробного дыхания на этом этапе приведен в табл. 9.2.
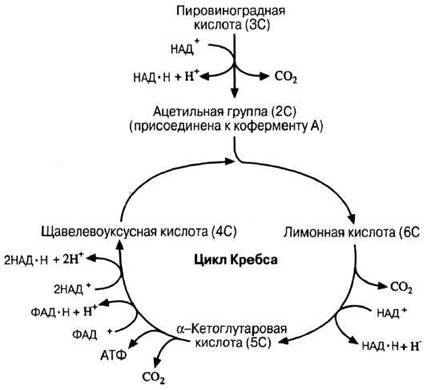
Рис. 9.6. Упрощенная схема цикла Кребса. Представлена также реакция, связывающая гликолиз с циклом Кребса (реакция между пировиноградной кислотой и коферментом А, в результате которой образуется ацетилкофермент А). Данная схема служит продолжением схемы рис. 9.5.
Таблица 9.2. Общий баланс окисления одной молекулы глюкозы при аэробном дыхании. Напоминаем, что для окисления одной молекулы глюкозы требуются два оборота цикла Кребса
СО2 |
АТФ |
НАД·Н + Н+ |
ФАД·Н + Н+ |
|
Гликолиз |
— |
2 |
2 |
— |
Пировиноградная кислота → Ацетил-КоА |
2 |
— |
2 |
— |
Цикл Кребса |
4 |
2 |
6 |
2 |
Итого: |
6СО2 |
4АТФ |
10(НАД · Н + Н+)* |
2(ФАД · Н+ Н+)* |
* Поступает в дыхательную цепь на кристах митохондрий. |
||||
Суммарное уравнение может быть записано в следующем виде:
![]()
Весь водород из молекулы глюкозы оказывается в конечном счете у переносчиков (НАД и ФАД). Весь углерод теряется в виде СО2. (Может вызвать удивление присутствие в этом уравнении шести молекул воды. Вода нужна в качестве источника кислорода в реакциях декарбоксилирования — именно такое происхождение имеет часть кислорода в СО2. Это, впрочем, деталь, которую можно и не учитывать.)
Дыхательная цепь и окислительное фосфорилирование
Водород на двух переносчиках (10 молекулах восстановленного НАД и двух молекулах восстановленного ФАД) направляется теперь к внутренней мембране митохондрий. Эта мембрана образует складки, так называемые кристы, увеличивающие площадь ее поверхности (рис. 9.12). Водород — это топливо. Мы уже отмечали, что при его окислении молекулярным кислородом образуется вода и выделяется энергия:
![]()
Часть этой энергии используется для синтеза АТФ из АДФ и неорганического фосфата при окислительном фосфорилировании (разд. 9.2.2.). Энергия не выделяется вся сразу в одной какой-нибудь реакции. Процесс разбит на ряд небольших этапов и среди них есть такие, на которых выделяется достаточно энергии для синтеза АТФ. Данная последовательность реакций известна как дыхательная цепь. В дыхательной цепи участвует ряд переносчиков водорода и электронов, заканчивается же она кислородом. Водород или электроны переходят от одного переносчика к другому, двигаясь в энергетическом смысле «вниз» до тех пор, пока на конечном этапе они не восстановят молекулярный кислород до воды. На каждом этапе выделяется некоторое количество энергии, причем в нескольких пунктах этот переход сопряжен с синтезом АТФ (эти пункты отмечены стрелками на рис. 9.7). В подписи к рис. 9.7 сказано о дыхательной цепи несколько более подробно. На конечном этапе действует медьсодержащий переносчик, называемый цитохромоксидазой. Цианид (или моноксид углерода) блокирует клеточное дыхание на этом этапе. Цианид связывается с медью, после чего кислород уже не может с ней соединиться.
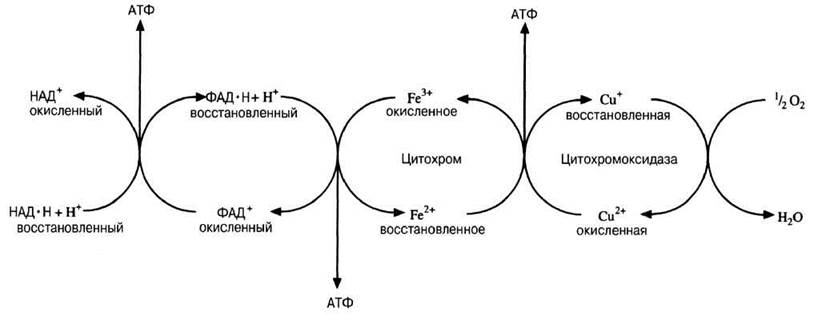
Рис. 9.7. Упрощенная схема дыхательной цепи. Водород переходит от восстановленного НАД к ФАД. Далее атомы водорода расщепляются на ионы водорода (Н+) и электроны. Электроны передаются от восстановленного ФАД железу (Fе), меди (Сu) и, наконец, кислороду, где они соединяются с ионами водорода, образуя воду. (Присоединение электрона есть восстановление, а его утрата — окисление; разд. 9.3.2.) Железо входит в состав гемогруппы белка, называемого цитохромом. Подобно гемоглобину, другому железосодержащему белку, цитохром окрашен (в розовый цвет). Медь входит в состав группы белков, носящих собирательное название цитохромоксидазы. Цитохромы переносят не водород, а электроны.
На рис. 9.7 видно, что на каждую молекулу восстановленного НАД, поступающую в дыхательную цепь, при переходе водорода или электронов к кислороду образуются 3 молекулы АТФ. Однако на каждую молекулу восстановленного ФАД образуется всего лишь две молекулы АТФ, потому что восстановленный ФАД поступает в дыхательную цепь на более низком энергетическом уровне.
Общий баланс для дыхательной цепи приведен в табл. 9.3.
Таблица 9.3. Баланс дыхательной цепи. Каждая молекула восстановленного НАД дает 3 молекулы АТФ и высвобождает водород, который соединяется с кислородом, образуя воду (Н2O). 10 молекул восстановленного НАД дают, следовательно, 30 молекул АТФ и 10 молекул воды. При этом используются 10 атомов кислорода, т. е. 5 его молекул. Каждая молекула восстановленного ФАД дает 2 молекулы АТФ
Поступает в дыхательную цепь |
Образуется |
Используется |
12Н2 в форме 10НАД · Н + Н+ и |
30АТФ + 10Н2О |
5O2 |
2ФАД · Н+ Н+ |
4АТФ + 2Н2O |
O2 |
Итого: |
34АТФ + 12Н2O* |
6O2 |
* Образуется 12H2O, но 6Н2О используются на более ранних этапах дыхания (см. уравнение в конце разд. 9.3.5), поэтому общий выход составляет 6Н2О. |
||
Суммарное уравнение для дыхательной цепи имеет вид:
![]()
Объединим два приведенных ниже уравнения, 1 и 2:
В результате получаем:
Итак, на каждую молекулу глюкозы, окисленную в процессе аэробного дыхания, образуется 38 молекул АТФ.
9.3. Какова роль кислорода в процессе дыхания?
Общая схема процесса аэробного дыхания приведена на рис. 9.8.
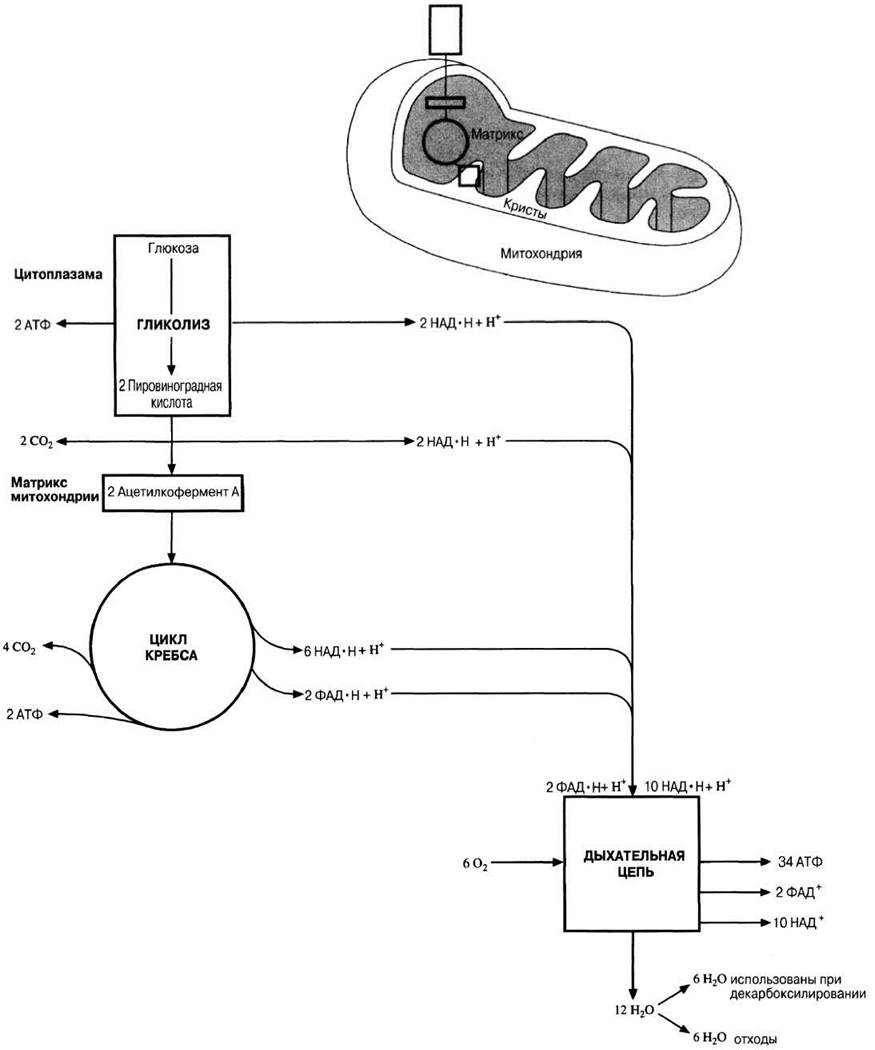
Рис. 9.8. Схема аэробного дыхания.
Окисление жирных кислот
Когда в качестве дыхательного субстрата используются липиды, они сначала гидролизуются до глицерола и жирных кислот, после чего от молекулы жирной кислоты последовательно отщепляются двууглеродные фрагменты, так что на каждом этапе эта длинная молекула укорачивается на два атома углерода. Двууглеродная ацетильная группа соединяется с коферментом А и образовавшийся ацетил-КоА вступает, как обычно, в цикл Кребса. Из каждой молекулы жирной кислоты извлекается большое количество энергии: при окислении стеариновой кислоты, например, выход АТФ составляет 147 молекул. Неудивительно поэтому, что жирные кислоты — важный источник энергии. Около половины обычных энергетических затрат сердечной мышцы, скелетных мышц (в покое), почек и печени покрывается именно за счет окисления жирных кислот.