БИОЛОГИЯ Том 2 - руководство по общей биологии - 2004
17. КООРДИНАЦИЯ И РЕГУЛЯЦИЯ У ЖИВОТНЫХ
17.5. Строение и функции рецепторов
17.5.3. Глаз
Глаз — это сенсорный орган, улавливающий электромагнитное излучение с определенными длинами волн (свет), которое испускается объектами или отражается от них в пределах некоторой части окружающего пространства — поля зрения, и преобразующий эти лучи в электрические импульсы. По зрительному нерву они поступают в головной мозг, где формируют воспринимаемый образ.
Глаз человека чувствителен только к излучению видимого спектра с длинами волн от 380 до 760 нм. Световая энергия испускается и поглощается дискретными порциями — квантами, или фотонами. Каждый квант волны видимого спектра способен вызвать фотохимическую реакцию в сенсорных клетках глаза — фоторецепторах.
Работа нашего глаза основана на тех же перечисленных ниже принципах, что и работа фотокамеры, а именно он; 1) контролирует количество света, попадающего внутрь; 2) фокусирует изображения предметов внешнего мира с помощью системы линз; 3) регистрирует изображение на чувствительной поверхности; 4) перерабатывает невидимое изображение во внутренний образ видимой картины мира.
Строение и функции человеческого глаза
Глазное яблоко по форме представляет собой примерно сферическую структуру диаметром около 24 мм и массой 6—8 г. Оно расположено в углублении черепа, называемом глазницей, и удерживается там четырьмя прямыми и двумя косыми мышцами, которые управляют его движениями. Основная часть глаза состоит из «вспомогательных структур», пропускающих свет к фоторецепторным клеткам, образующим самый внутренний слой сферы — сетчатку. Она окружена сосудистой оболочкой, переходящей спереди в ресничное тело и радужку со зрачком. Наружный слой глазного яблока — фиброзная оболочка — подразделяется на роговицу и склеру (рис. 17.33). Непосредственно за зрачком расположен хрусталик. Внутренняя часть сферы занята стекловидным телом и так называемой водянистой влагой, которые создают внутриглазное давление, близкое к 3,3 кПа.
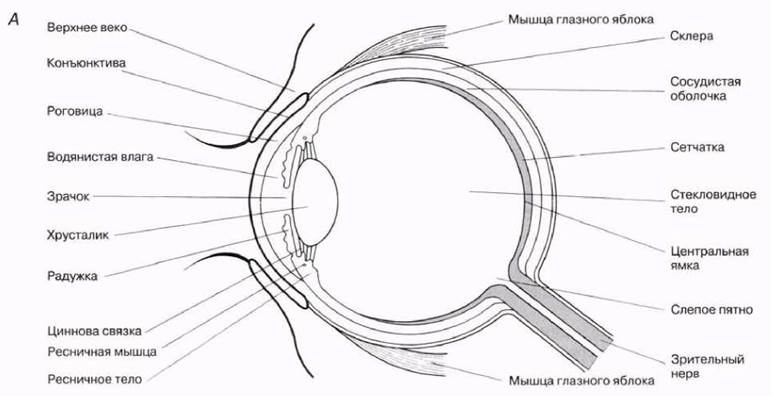

Рис. 17.33. Строение глаза человека: А —схема вертикального продольного среза; Б — препарат, на основе которого построена схема.
Строение человеческого глаза показано на рис. 17.33, а функции различных его частей вкратце перечислены ниже.
Склера — плотная, богатая коллагеновыми волокнами оболочка белого цвета, защищающая глаз от повреждений и поддерживающая его форму.
Роговица — прозрачная передняя сторона склеры; благодаря искривленной поверхности действует как главная светопреломляющая структура, направляющая световые лучи на сетчатку.
Конъюнктива — тонкий прозрачный соединительнотканный слой клеток, защищающий роговицу и переходящий в эпителий внутренней поверхности век.
Веко — кожная складка, защищающая путем рефлекторных движений переднюю поверхность глазного яблока от механических и химических повреждений, а сетчатку — от слишком яркого света.
Сосудистая оболочка — слой, пронизанный кровеносными сосудами, питающими сетчатку, и выстланный изнутри черным пигментным эпителием, предотвращающим отражение света внутри глаза.
Ресничное (цилиарное) тело — место соединения склеры и роговицы; содержит эпителиальные клетки, кровеносные сосуды и ресничную мышцу.
Ресничная мышца — кольцо, состоящее из гладких мышечных волокон, кольцевых и радиальных, которые изменяют кривизну хрусталика в процессе аккомодации.
Цилиарная (циннова) связка — соединяет хрусталик с ресничным телом.
Хрусталик — прозрачная упругая двояковыпуклая линза; обеспечивает тонкую фокусировку лучей света на сетчатке за счет изменения своей кривизны и разделяет камеры, заполненные водянистой влагой и стекловидным телом.
Водянистая влага — прозрачный солевой раствор, секретируемый ресничным телом, заполняющий переднюю и заднюю камеры глаза между роговицей и хрусталиком; переходит в кровь через шлеммов канал.
Радужка — кольцевая диафрагма, содержащая пигмент, определяющий цвет глаз; разделяет пространство, заполненное водянистой влагой, на переднюю и заднюю камеры и регулирует количество проникающего в глаз света.
Зрачок — центральное отверстие радужки, пропускающее свет внутрь глаза.
Стекловидное тело — прозрачная желеобразная масса, окруженная мембраной, заполняющая изнутри глазное яблоко и поддерживающая его форму.
Центральная ямка — самая сильная по разрешающей способности (остроте зрения) часть сетчатки диаметром менее 0,5 мм, содержащая только колбочки. Здесь фокусируется основная часть световых лучей.
Зрительный нерв — пучок аксонов, проводящий импульсы от сетчатки в головной мозг.
Слепое пятно — место отхождения зрительного нерва от сетчатки (пятно диаметром примерно 1,7 мм); оно не содержит ни палочек, ни колбочек) и поэтому не обладает светочувствительностью.
17.6. Перечислите по порядку структуры, через которые проходит свет, прежде чем достигнет сетчатки.
Аккомодация
Аккомодация — это рефлекторный механизм, посредством которого световые лучи, исходящие от объекта, фокусируются на сетчатке. Он включает в себя два процесса, каждый из которых мы рассмотрим по отдельности.
ЗРАЧКОВЫЕ РЕФЛЕКСЫ. При ярком свете кольцевая мышца радужки (сфинктер зрачка) сокращается, а радиальная (расширитель зрачка) расслабляется. В результате зрачок сужается, уменьшая поток световых лучей внутрь глаза и тем самым предотвращая повреждение сетчатки (рис. 17.34). При тусклом свете, наоборот, радиальная мускулатура сокращается, а кольцевая расслабляется, и зрачок расширяется. Дополнительное преимущество сужения зрачка — чисто оптическое явление, называемое увеличением глубины резкости: чем уже пропускающее свет отверстие, тем четче видны через него объекты.
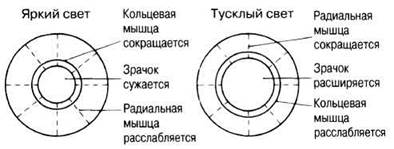
Рис. 17.34. Реакции радужной оболочки (и тем самым зрачка) на изменения освещенности.
ПРЕЛОМЛЕНИЕ (РЕФРАКЦИЯ) СВЕТА. Световые лучи от удаленных объектов (на расстоянии более 6 м) идут в глаз практически параллельно, а от более близких — расходящимся пучком. В обоих случаях для получения четкого изображения на сетчатке их надо преломить и сфокусировать на ней, причем для близких предметов преломление должно быть более сильным. Нормальный глаз способен точно фокусировать свет от объектов, находящихся на расстоянии примерно от 25 см до «бесконечности». Преломление света происходит при переходе его из одной среды в другую, имеющую иной коэффициент преломления, в частности на границе воздух—роговица и у поверхностей хрусталика.
Кривизна роговицы постоянна, поэтому рефракция здесь зависит только от угла падения света на роговицу, а этот угол в свою очередь зависит от удаленности предмета. Роговица наиболее сильно преломляет свет, а функция хрусталика состоит в окончательной «наводке на фокус». Форма хрусталика регулируется ресничной мышцей: от степени ее сокращения зависит натяжение связки, поддерживающей хрусталик. Последняя воздействует на упругий хрусталик и изменяет его форму (радиус кривизны), а, следовательно, и угол преломления света. Когда радиус уменьшается, ресничное тело приближается к хрусталику, натяжение ресничного пояска, на котором он укреплен, ослабевает, и хрусталик из-за своих упругих свойств становится более выпуклым. Это усиливает преломление света, и глаз «настраивается» на рассматривание близких объектов. Расслабление ресничной мышцы приводит к упругому удалению ресничного тела от хрусталика, напряжению ресничного пояска и растягиванию хрусталика. Он уплощается, преломление света ослабевает, и устанавливается новое фокусное расстояние, «настроенное» на дальние объекты. Эти изменения обобщены в табл. 17.7 и на рис. 17.35.
Таблица 17.7. Связь между структурами, участвующими в изменении формы хрусталика, и степенью преломления света
Ресничная мышца |
Циннова связка |
Кривизна хрусталика |
Преломление света |
Сокращена (объект близко) |
Не натянута |
Увеличена (хрусталик более выпуклый) |
Усилено |
Расслаблена (объект далеко) |
Натянута |
Уменьшена |
Ослаблено |

Рис. 17.35. Аккомодация при восприятии объектов, находящихся на разных расстояниях от глаза. А. Вид глаза сбоку. Б. Вид глаза спереди.
Изображение, построенное на сетчатке, по законам оптики является перевернутым и обратным, однако мы без труда ориентируемся в окружающем мире, поскольку головной мозг (зрительная кора), обрабатывая сенсорную информацию, позволяет нам воспринимать увиденное, «как надо».
Строение сетчатки
Фоторецепторные клетки сетчатки (палочки и колбочки) направлены своими светочувствительными частями к сосудистой оболочке, а со стороны стекловидного тела перекрыты телами и отростками нейронов, связывающих их с головным мозгом (рис. 17.36).
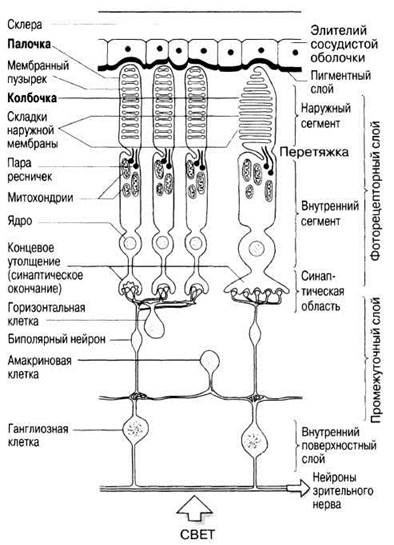
Рис. 17.36. Схема строения сетчатки глаза с деталями ультраструктуры палочек и колбочек. Показаны связи между сенсорными клетками и нейронами зрительного нерва. Лучи света должны пройти через слои ганглиозных, амакриновых и биполярных клеток, прежде чем они достигнут палочек и колбочек.
Сетчатка состоит из трех слоев, различающихся по типам образующих их клеток. Самый наружный фоторецепторный слой состоит из светочувствительных клеток — палочек и колбочек, частично погруженных в пигментный эпителий сосудистой оболочки. Затем идет промежуточный слой, содержащий биполярные нейроны, синаптически связывающие фоторецепторы с клетками третьего слоя, а также горизонтальные и амакриновые клетки, обеспечивающие так называемое латеральное торможение (см. ниже). Третий — внутренний поверхностный слой — образован ганглиозными клетками, дендриты которых контактируют с биполярными нейронами, а аксоны формируют зрительный нерв.
Строение и функция палочек и колбочек
Палочки и колбочки очень сходны по своему строению (рис. 17.36). В тех и других фоточувствительные (зрительные) пигменты находятся на наружной поверхности собранных в стопки мембран так называемого наружного сегмента клетки; и те и другие состоят из четырех участков, строение и функция которых кратко описаны ниже.
НАРУЖНЫЙ СЕГМЕНТ. Это тот светочувствительный участок, где световая энергия преобразуется в генераторный потенциал. Весь сегмент состоит из стопки уплощенных мембранных пузырьков, содержащих фотосинтетические пигменты. В палочке они представлены 600—1000 таких пузырьков, окруженных наружной мембраной, а в колбочке — складками самой наружной мембраны (их число меньше). Наружный сегмент палочек цилиндрический, а колбочек — конический.
ПЕРЕТЯЖКА. Здесь наружный сегмент почти полностью отделен от внутреннего впячиванием наружной мембраны. Связь между двумя сегментами осуществляется через цитоплазму и пару ресничек, переходящих из одного сегмента в другой. Каждая ресничка состоит только из девяти периферических групп микротрубочек (центральная их пара, типичная для таких органелл, отсутствует) и не выполняет двигательных функций.
ВНУТРЕННИЙ СЕГМЕНТ. Это область активного метаболизма; она заполнена митохондриями, поставляющими энергию для процессов зрения, и полирибосомами для синтеза белков, участвующих в образовании мембранных пузырьков и зрительных пигментов. В этом же участке расположено ядро.
СИНАПТИЧЕСКАЯ ОБЛАСТЬ. Здесь фоторецепторные клетки образуют синапсы с биполярными клетками. Некоторые биполярные клетки связаны синапсами сразу с несколькими палочками, что обеспечивает конвергенцию раздражения, которая, как уже говорилось, повышает светочувствительность глаза, но снижает остроту зрения (разд. 17.4.2). Другие биполярные клетки связывают одну колбочку с одной ганглиозной клеткой, поэтому острота «колбочкового зрения» выше, а чувствительность ниже. Горизонтальные и амакриновые клетки связывают между собой по несколько палочек и колбочек. Благодаря этим клеткам зрительная информация еще до выхода из сетчатки подвергается определенной обработке, в частности, они участвуют в латеральном торможении (см. ниже).
Различия между палочками и колбочками
Палочек в сетчатке содержится больше, чем колбочек (соответственно 120 млн. и 6 млн.). Распределение этих фоторецепторов в сетчатке тоже неодинаково. Палочки равномерно распределены по всей сетчатке, кроме центральной ямки, где преобладают колбочки. Так как в центральной ямке колбочки плотно упакованы (50 000 на 1 мм), этот участок отличается высокой остротой зрения (см. ниже).
Палочки (в своей совокупности) гораздо чувствительнее к свету, чем колбочки, и реагируют даже на небольшое его количество. Они содержат только один зрительный пигмент, поэтому не способны различать цвета и служат нам главным образом в темное время суток. В каждой колбочке содержится один из трех зрительных пигментов, взаимодействие которых обеспечивает цветовое зрение. Однако используются эти фоторецепторы в связи со своей низкой чувствительностью главным образом в дневное время.
Палочковое зрение отличается меньшей остротой, поскольку палочки упакованы не так плотно, как колбочки в центральной ямке, и сигналы от них подвергаются конвергенции, но именно это обеспечивает высокую чувствительность, необходимую для ночного зрения (см. ниже).
Чувствительность и острота зрения
Острота зрения — это его разрешающая способность, т. е. степень различения мелких деталей объекта. Так, если один участок сетчатки может видеть две близко расположенные точки как две, а для другого участка они сливаются в одну, то в первом случае острота зрения выше. Максимальна она в центральной ямке, которая обычно воспринимает центральную часть нашего поля зрения, и понижается к его краям. Поэтому, внимательно рассматривая объект, мы переводит взгляд с одной его части на другую, располагая их поочередно перед центральной ямкой. Здесь находятся только колбочки, причем около 90% их общего числа. Многие из них синаптически связаны всего с одним биполярным нейроном (рис. 17.36), который в свою очередь связан с единственной ганглиозной клеткой. Это соотношение 1:1 обеспечивает максимальную теоретическую остроту зрения, поскольку каждая часть изображения воспринимается особой клеткой, а значит, передаваемая ей информация не сливается и не «размывается» потоком других зрительных сигналов. Острота зрения повышается с увеличением числа колбочек на единицу площади (как на фотографии в газете: чем больше образующих ее точек, тем четче картинка). Колбочки, удаленные от центральной ямки (а некоторые и в ней), связаны с двумя-тремя биполярными нейронами, поэтому к периферии сетчатки острота зрения снижается.
Примерно на 120 млн. палочек и 6 млн. колбочек в сетчатке каждого глаза приходится всего 1,2 млн. ганглиозных клеток, следовательно, в них происходит конвергенция сигналов (разд. 17.4.2) в среднем от 105 фоторецепторов. Для палочек степень этой конвергенции гораздо выше, чем для колбочек, поэтому обеспечиваемая ими острота зрения намного ниже, но выше его чувствительность (см. вопрос 17.7 ниже). При слабом освещении хуже всего воспринимает свет центральная ямка, где палочек вообще нет, а лучше всего — периферия сетчатки, где они преобладают. В этом можно убедиться, глядя ясной ночью на небо — по краям поля зрения мы различаем гораздо больше звезд, чем в его центре. Когда света мало, работают только палочки, поэтому острота зрения падает (воспринимаемое изображение становится размытым), а способность различать цвета исчезает.
17.7. Объясните, почему синаптическая конвергенция повышает чувствительность зрения.
17.8. Объясните, почему ночью мы лучше различаем объекты, если смотрим на них не прямо, а «краем» глаза?
Механизм фоторецепции
Палочки содержат светочувствительный пигмент родопсин (зрительный пурпур), находящийся на наружной поверхности мембранных дисков. Родопсин представляет собой сложную молекулу, образованную белком скотопсином (олеином) и обратимо связанной с ним поглощающей свет небольшой молекулой каротиноида ретиналя. Последний является производным витамина А и существует в двух (цис- и транс-) изомерных формах, переходящих друг в друга в зависимости от условий освещения (рис. 17.37).
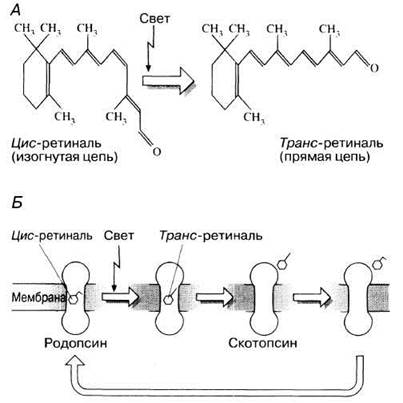
Рис. 17.37. А. Переход цис-ретиналя в транс-ретиналь под действием света. Б. Выцветание и регенерация родопсина.
Известно, что воздействие одного фотона света приводит к превращению цис-молекулы ретиналя в его транс-форму. При этом происходит отделение ретиналя от скотопсина (процесс, называемый выцветанием пигмента):
![]()
В темноте родопсин сразу же ресинтезируется: вначале транс-ретиналь превращается в цис-форму и затем последний соединяется со скотопсином (рис. 17.37, Б). Этот процесс составляет химическую основу темновой адаптации и в полной темноте примерно за 30 мин происходит во всех палочках, тем самым возвращая глазу его максимальную светочувствительность.
Каким же образом химическое разложение родопсина на свету приводит к генерированию потенциала действия? Это связано с изменением мембран наружного и внутреннего сегментов палочек (рис. 17.38). Во внутреннем сегменте палочки функционирует натриевый насос, непрерывно выкачивающий из клетки ионы натрия. В темноте мембрана наружного сегмента для них проницаема, и они диффундируют назад, в результате чего поддерживается стабильный мембранный потенциал (—40 мВ, а не —70 мВ, как у большинства клеток). На свету, однако, проницаемость снижается, а действие натриевого насоса продолжается, приводя к росту отрицательного заряда внутри палочки (рис. 17.38). В конечном итоге происходит гиперполяризация ее мембраны. Эта ситуация прямо противоположна тому, что обычно происходит в других рецепторных клетках, у которых вызываемый сигналом генераторный потенциал обусловлен, напротив, деполяризацией мембраны. Гиперполяризация палочек замедляет высвобождение из них возбуждающего синапсы медиатора, который, таким образом, наиболее активно выделяется в темноте. В результате происходит гиперполяризация связанных с фоторецепторами биполярных нейронов, однако ганглиозные клетки, с которыми контактируют эти нейроны, в ответ на сигнал от них генерируют потенциал действия.
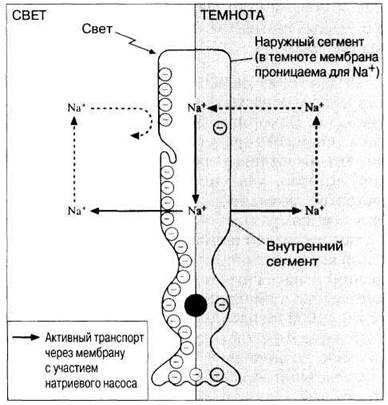
Рис. 17.38. Схема строения палочки, иллюстрирующая предполагаемые изменения проницаемости наружного сегмента для ионов натрия под действием света. Отрицательные заряды (кружочки с минусом внутри) на правой стороне палочки соответствуют потенциалу покоя, на левой стороне — гиперполяризации.
Роль горизонтальных и амакриновых клеток
Горизонтальные клетки, каждая из которых стимулируется несколькими палочками и синаптически связана с несколькими биполярными нейронами (рис. 17.36), обеспечивают феномен так называемого латерального (бокового) торможения, повышающего как чувствительность, так и остроту зрения. Упрощенно говоря, если эти клетки одновременно получают сигналы одинаковой интенсивности от двух соседних палочек, то они их «взаимно гасят», т. е. тормозят дальнейшее проведение. В результате эффективными оказываются только сигналы от неодинаково возбужденных рецепторов, а это повышает контрастность изображения — подчеркивается разница между сильно и слабо освещенными участками поля зрения, что позволяет лучше различать, например, контуры объектов. Амакриновые клетки, каждая из которых стимулируется несколькими биполярными нейронами и синаптически связана с несколькими ганглиозными клетками, передают информацию об изменениях уровня освещенности.
Цветовое зрение
Человеческий глаз поглощает свет всех длин волн видимой части спектра, воспринимая их в виде шести разных цветов, каждый из которых примерно соответствует определенному участку спектра (табл. 17.8). Цвета как таковые в природе не существуют — это «изобретение» головного мозга. Различение их обусловлено наличием трех типов колбочек, которые содержат разные пигменты, поглощающие волны в своем диапазоне длин. Таким образом, в зависимости от максимальной спектральной чувствительности выделяют «красные», «зеленые» и «синие» колбочки. Эксперименты показали, что любые оттенки можно получить путем сочетания этих трех чистых цветов.
17.9. Используя табл. 17.8, предположите, какого цвета объект вы увидите, если смотрите на него, когда один ваш глаз закрыт зеленым фильтром (средняя длина волны 530 нм), а другой - красным (средняя длина волны 620 нм).
Таблица 17.8. Цвета видимого спектра и примерные диапазоны соответствующих им длин волн
Цвет |
Длины волн, нм |
Красный |
Более 620 |
Оранжевый |
590-620 |
Желтый |
570-590 |
Зеленый |
500-570 |
Синий |
440-500 |
Фиолетовый |
Менее 440 |
Цветовое зрение объясняют с позиций наиболее популярной трихроматической теории, согласно которой воспринимаемый цвет зависит от степени стимуляции каждого типа колбочек. Например, одинаковая стимуляция всех колбочек вызывает ощущение белого цвета. Рис. 17.39 демонстрирует чувствительность трех типов колбочек к разным длинам волн. Очевидно, что, хотя они и называются «красными», «синими» и «зелеными», каждая воспринимает не только «свой» цвет, но и некоторые другие, только слабее, причем области чувствительности разных типов фоторецепторов частично перекрываются. Следовательно, видимый цвет определяется просто разной степенью стимуляции каждого типа колбочек. Например, оранжевые лучи примерно одинаково возбуждают зеленые и красные колбочки, синие сильно стимулируют синие рецепторы и слабо — зеленые, а зеленые лучи активируют все три типа колбочек.
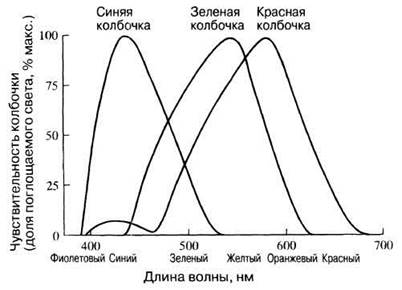
Рис. 17.39. Чувствительность трех типов колбочек к свету различных длин волн (цветам спектра).
Трихроматическая теория объясняет большую часть экспериментальных данных по цветовому зрению, однако некоторые факты до сих пор остаются неясными. Их обсуждение выходит за рамки настоящей книги, однако они свидетельствуют о необходимости дальнейших теоретических разработок в этой области.
Первичное различение цветов происходит в сетчатке, однако окончательное формирование воспринимаемого оттенка требует интерпретации сенсорных сигналов головным мозгом.
Цветовая слепота. Полное отсутствие или недостаток колбочек какого-либо типа ведет к различным формам цветовой слепоты или аномалиям цветоощущения, т. е. неспособности различать определенные цвета. Например, люди, у которых отсутствуют «красные» или «зеленые» колбочки, не различают красный и зеленый цвета, а те, у кого имеется недостаточное число либо тех, либо других колбочек, плохо различают «ненасыщенные» оттенки этих цветов. Для выявления дефектов цветового зрения применяют специальные тестовые таблицы (например, таблицы Исихары), составленные из разноцветных точек. На некоторых таблицах из этих точек составлены цифры. Человек с нормальным цветовым зрением легко различает эти цифры, а лица с нарушенным цветоощущением видят другое число или вообще не видят никакой цифры. Цветовая слепота передается по наследству как рецессивный признак, сцепленный с полом. Он обусловлен генными дефектами Х-хромосомы, поэтому им чаще всего страдают мужчины: около 2% их не видят красного цвета (протанопы), а 6% — зеленого (дейтеранопы), тогда как оба типа этих аномалий встречаются только у 0,4% женщин.
Бинокулярное и стереоскопическое зрение
Бинокулярное зрение — это восприятие объекта одновременно двумя глазами. При этом их поля зрения перекрываются, и изображение фокусируется одновременно в двух центральных ямках сетчатки. Поскольку каждый глаз смотрит на объект под своим углом, два изображения несколько различаются, но интерпретируются зрительной корой головного мозга как единый образ. Бинокулярное зрение лежит в основе стереоскопического, т. е. объемного, зрения; в этом случае мы различаем относительную удаленность от нас разных видимых деталей объекта, иными словами, мы воспринимаем глубину пространства.
Чем ближе друг к другу расположены глаза, тем сильнее перекрываются их поля зрения и тем больше область стереоскопического зрения. Например, у человека общее поле зрения обоих глаз охватывает угол 180°, а стереоскопическое — 140°. У лошади глаза расположены по бокам головы, и объемно она воспринимает только удаленные объекты в узкой полосе прямо перед собой. Чтобы рассмотреть близкий предмет, лошадь поворачивает голову и пользуется монокулярным зрением, а это дает плоский зрительный образ, как на киноэкране. Для хорошего стереоскопического зрения необходимы глаза, направленные вперед, с центральными ямками, лежащими посередине их полей, что обеспечивает большую остроту зрения. В этом случае стереоскопическое зрение позволяет получать более точное представление о размерах и форме предмета и о расстоянии до него. Стереоскопическое зрение свойственно главным образом хищникам, которым оно жизненно необходимо, если они ловят добычу, внезапно набрасываясь на нее или пикируя с высоты, как это делают представители семейства кошачьих, ястребы и орлы. У животных, чья стратегия выживания связана с бегством от врага, глаза, как правило, расположены по бокам головы — это расширяет общее поле зрения, сужая его бинокулярную область (у кролика, например, они соответственно равны 360° и 26°): им важнее как можно раньше заметить опасность и броситься «без оглядки» в противоположную от нее сторону.
Анализ изображений, получаемых на сетчатке при стереоскопическом зрении, осуществляется в двух симметричных участках, составляющих зрительную кору.
Роль головного мозга
Нервные импульсы, возникающие в сетчатке, примерно по миллиону волокон зрительного нерва направляются в первичную зрительную зону зрительной коры, находящейся на затылочной доле больших полушарий головного мозга (рис. 17.26). Здесь каждому фрагменту сетчатки, включающему, вероятно, всего несколько палочек и(или) колбочек, соответствует свой участок, обрабатывающий сигналы таким образом, что мы «видим». Однако, что мы видим приобретает смысл только после обмена сигналами с другими зонами коры, и прежде всего с височными долями, где хранится предшествующая зрительная информация и где она используется для анализа и интерпретации текущих зрительных сигналов (разд. 17.2.4). У человека аксоны от левой стороны сетчатки обоих глаз (она по законам оптики воспринимает правую часть поля зрения) направляются в левое полушарие, а от правой половины (левая часть поля зрения) — в правое. Аксоны, отходящие от медиальных (ближайших к носу) половин сетчатки пересекаются; место их пересечения хорошо заметно на нижней поверхности головного мозга в виде крестовины, образуемой сходящимися зрительными нервами; это — так называемый зрительный перекрест или хиазма (рис. 17.40). Примерно 20% волокон зрительного нерва не доходят до коры больших полушарий и оканчиваются в среднем мозге, где участвуют в формировании рефлексов, связанных с изменением размеров зрачка и движениями глаз.

Рис. 17.40. Схема зрительных путей человека. Вид с нижней стороны головного мозга.