БИОЛОГИЯ Том 2 - руководство по общей биологии - 2004
13. ТРАНСПОРТ У РАСТЕНИЙ
13.8. Транслокация органических веществ по флоэме
13.8.4. Механизм транслокации веществ по флоэме
В разд. 13.8.1 и 13.8.3 были перечислены факты, которые должны быть учтены в любой гипотезе, объясняющей этот транспорт; суть этих фактов сводится к следующему: большие количества вещества относительно быстро перемещаются по очень тонким ситовидным трубкам. Внутри трубок существуют очевидные препятствия — ситовидные пластинки; кроме того, есть и другие структурные особенности, в частности флоэмный белок, о роли которого ничего не известно. Если добавить к этому, что вся система довольно хрупкая и легко повреждается при всяком постороннем вмешательстве, то можно понять те трудности, с которыми столкнулись ученые, выясняющие механизм флоэмной транслокации.
Многие сейчас считают, что по ситовидным трубкам движется объемный поток раствора. Диффузия — слишком медленный процесс, чтобы обеспечить наблюдаемую скорость транспорта. Ниже мы перечислим данные, свидетельствующие о том, что по флоэме идет объемный поток вещества.
1. Когда флоэму перерезают, из разреза начинает вытекать сок; очевидно, он движется по принципу объемного потока. Иногда это используют в практических целях для получения сахара. Так, например, сахарная пальма «плачет», выделяя ежедневно по 10 л сахаристого сока.
2. Продолжительное выделение раствора сахарозы из хоботка тли в описанном выше (разд. 13.8.3) эксперименте говорит о наличии в ситовидных трубках гидростатического давления.
3. С флоэмным током по растению переносятся некоторые вирусы. Вирусы не способны к активному движению и, будучи нерастворимыми, не могут диффундировать. Единственное объяснение их перемещения — движение в объемном потоке жидкости.
Гипотезы объемного потока Мюнха и тока под давлением
В 1930 г. Мюнх предложил чисто физическую гипотезу, объясняющую формирование объемного потока в ситовидных трубках. Эта гипотеза проиллюстрирована с помощью модели на рис. 13.26.

Рис. 13.26. Физическая модель, которая иллюстрирует гипотезу объемного потока Мюнха, объясняющую передвижение растворимых веществ по флоэме. Моделируемые части живого растения: А — источник ассимилятов, например лист; В — флоэма; С — пункт доставки веществ, например корень, меристемы, плоды; D — ксилема, апопласт и межклетники.
В этой модели вода исходно стремится переходить путем осмоса в сосуды А и С, но для А это стремление сильнее, поскольку раствор в А более концентрирован, чем в С. Поступление воды в А повышает в замкнутой системе А-В-С гидростатическое давление, заставляющее воду выходить из С. В трубке В возникает объемный поток, движущийся вдоль градиента давления. Между А и С существует также осмотический градиент. В конечном итоге по мере разбавления раствора в А и перекачки растворенного вещества в С система уравновешивается.
Эту модель можно применить к живым растениям. Сосуд А соответствует листьям. Образование в них сахара путем фотосинтеза делает осмотический (ψ0), а значит, и водный потенциалы более отрицательными. Вода, поступающая к листьям по ксилеме (D), проникает в мезофилл путем осмоса, повышая его гидростатический потенциал (ψг). Одновременно сахара расходуются в своих «конечных пунктах», например в корнях (С), для самых разных целей, в том числе для дыхания и синтеза целлюлозы. Это повышает там осмотический потенциал (ψ0), т. е. делает его менее отрицательным, тормозя осмос. В результате формируется гидростатический градиент, снижающийся от листьев к корням, или, в общем случае, от места образования ассимилятов к пунктам их потребления, что приводит к объемному потоку жидкости между ними. В живом растении равновесие никогда не достигается, поскольку растворимые вещества непрерывно расходуются тканями-потребителями (С) и образуются в ассимилирующих тканях (А).
Гипотеза Мюнха чисто физическая, и не объясняет, почему ситовидные трубки должны оставаться живыми и метаболически активными. Она не объясняет также, каким образом клетки мезофилла листа способны загружать ситовидные трубки ассимилятами против осмотического градиента: известно, что у флоэмы ψ0 более отрицательный, чем у фотосинтезирующей ткани. С учетом этого гипотеза Мюнха была впоследствии дополнена — в нее включили механизм активной загрузки растворенных веществ в ситовидные трубки. Он подразумевает, что осмотический и гидростатический градиенты начинаются не в фотосинтезирующем мезофилле, а непосредственно во флоэме. Кроме того, полагают, что разгрузка флоэмы на уровне потребителей — тоже активный процесс. Такая современная версия гипотезы Мюнха называется гипотезой тока под давлением.
Загрузка ситовидных трубок
Было показано, что концентрация сахарозы в ситовидных трубках листьев обычно составляет от 10 до 30%, тогда как в фотосинтезирующих клетках она держится на уровне 0,5%. Следовательно, как мы уже отмечали, загрузка флоэмы идет против осмотического градиента. Этот механизм активно изучался в последние годы. Сначала растворенные органические вещества должны переместиться из хлоропластов во флоэму; этот путь составляет не более 3 мм, причем вещества могут двигаться и по симпласту, и по апопласту. Симпластный путь предполагает перемещение веществ по плазмодесмам, а апопластный — через клеточные стенки посредством диффузии или объемного потока в транспирационном паре.
В 1968 г. Гуннинг и его сотрудники описали новый тип модифицированных клеток-спутниц — передаточные клетки. Как видно из рис. 13.27, эти клетки примыкают к ситовидным трубкам. В результате дополнительного неравномерного утолщения клеточные стенки у них образуют многочисленные внутренние выступы, что почти в десять раз увеличивает площадь выстилающей такую стенку плазмалеммы. Как полагают, такое видоизменение клеток связано с необходимостью активно поглощать растворенные вещества из соседних клеток. Энергию для всех активных процессов дают многочисленные митохондрии передаточных клеток. Передаточные клетки обнаружены не у всех растений, но они обычны у бобовых и некоторых других семейств. Впрочем, считают, что активный транспорт происходит и в их отсутствие.

Рис. 13.27, Поперечный срез мелкой жилки листа крестовника обыкновенного (Senecio vulgaris). Во флоэме (верхняя половина снимка) видны шесть Ю1еток, окружающих две ситовидные трубки. Это четыре клетки-спутницы (КС) с плотной цитоплазмой, преобразованные в передаточные клетки, и две клетки флоэмной паренхимы (ФП) с более прозрачной цитоплазмой и выростами клеточной стенки только со стороны, прилегающей к ситовидным трубкам. Последние обычно соединены с клетками-спутницами плазмодесмами, а флоэмная паренхима образует симпластную связь с проводящими элементами очень редко. В нижней части снимка посередине видны два ксилемных сосуда, а по бокам от них — две крупные клетки обкладки проводящего пучка. Стрелки указывают возможные пути движения растворенных веществ в ситовидные трубки, включая апопластный транспорт некоторых компонентов ксилемного сока. Увеличение в 6560 раз.
Как полагают, загрузку сахарозы (и ряда других компонентов раствора, в том числе аминокислот, фосфатов, ионов калия и аммония) в клетки-спутницы осуществляют особые белки-переносчики, локализованные в плазмалемме этих клеток. Образуемые белками системы переноса сходны с аналогичными системами животных и бактериальных клеток, где транспорт органических молекул сопряжен с транспортом прогонов (Н+). Протоны выкачиваются из клетки переносчиком, использующим для этого энергию АТФ (рис. 13.28). В результате формируется градиент концентрации Н+, генерирующий потенциальную энергию. Протоны быстро диффундируют назад в клетку с участием других белков-перепосчиков, которые одновременно пропускают внутрь молекулы сахарозы или других органических веществ (котранспорт по типу симпорта). Таким образом, движущая сила этого процесса — формирование протонного градиента через плазмалемму с пониженным pH (более высокой концентрацией Н+) вне клетки.
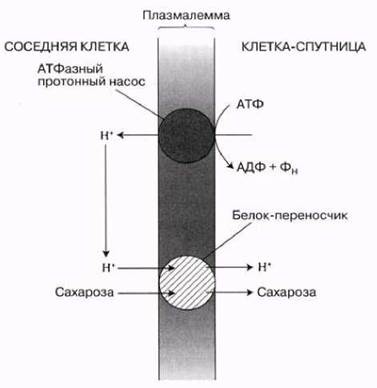
Рис. 13.28. Загрузка клетки-спутницы сахарозой. Ионы Н+ (протоны) выкачиваются из этой клетки протонным насосом. Он находится в плазмалемме, требует для своей работы энергии АТФ и сам обладает АТФазной активностью. Протоны возвращаются в клетку по механизму облегченной диффузии: особый белок-переносчик пропускает их туда одновременно с молекулами сахарозы (симпорт).
В результате активного транспорта веществ в клетки-спутницы осмотический потенциал в них сильно понижается. Это стимулирует поступление в них воды путем осмоса, повышение давления и движение растворов (в том числе и раствора сахарозы) по механизму объемного потока через многочисленные плазмодесмы, связывающие клетки-спутницы с ситовидными трубками. Таким образом высокое давление и объемный поток создаются в ситовидных трубках, а не в клетках мезофилла листа, как предполагал Мюнх. Возможно также, что механизм активного транспорта через плазмодесмы из клеток-спутниц в ситовидные трубки точно такой же, как и механизм, обеспечивающий поступление сахарозы в клетки-спутницы.
При разгрузке ситовидных трубок растворенные вещества, вероятно, уходят из них в передаточные клетки по плазмодесмам. В результате водный потенциал трубок повышается (становится менее отрицательным). Это тормозит потенциальный осмос извне и поддерживает гидростатический градиент между местами загрузки и разгрузки флоэмы. Полагают, что последний процесс может идти и через плазмалемму ситовидных трубок в их клеточные стенки и далее апопластным путем.
Критическая оценка гипотезы тока под давлением
1. Согласно этой гипотезе, в ситовидных трубках формируется объемный поток, что соответствует фактам (см. данные, приведенные выше).
2. Гипотеза предполагает существование осмотического градиента и высокого давления во флоэме. Это показано для целого ряда растений.
Гидростатический градиент, необходимый для движения растворов по ситовидным трубкам с наблюдаемой скоростью, должен быть относительно крутым. Даже если все ситовидные поры открыты, теоретический расчет дает величину градиента, равную 13 кПа/м, которая до недавнего времени считалась вряд ли достижимой. Непосредственное измерение флоэмного давления — задача очень сложная, однако в последние годы она все же была решена. Зарегистрированы его уровни в пределах 1000—2000 кПа с градиентом до 20 кПа/м. Таким образом, и здесь гипотеза подтвердилась.
3. Гипотезу тока под давлением критиковали за то, что она не объясняет, почему ситовидные трубки остаются живыми в отличие от мертвых ксилемных сосудов.
Однако только у живых клеток есть плазмалемма, необходимая для предотвращения утечки сахарозы во внешнюю среду. Судя по результатам недавних исследований, интенсивность обмена веществ в ситовидных трубках низка, а это предполагает пассивное движение через них растворов, что полностью соответствует гипотезе тока под давлением.
4. Считают, что ситовидные пластинки, несмотря на создаваемое ими сопротивление потоку, необходимы, поскольку они обеспечивают прочность ситовидных трубок, не позволяя им раздуваться, трескаться или лопаться под действием высокого внутреннего давления.
5. У содержимого ситовидных трубок pH близок к 7,5—8 (слабощелочная среда). Это можно объяснить тем, что протоны, выкачиваются из них, как это было описано выше. Присутствие протонного насоса объясняет и высокую концентрацию АТФ, отмечаемую в ситовидных трубках. Общая схема движения сахарозы по растению приведена на рис. 13.29.
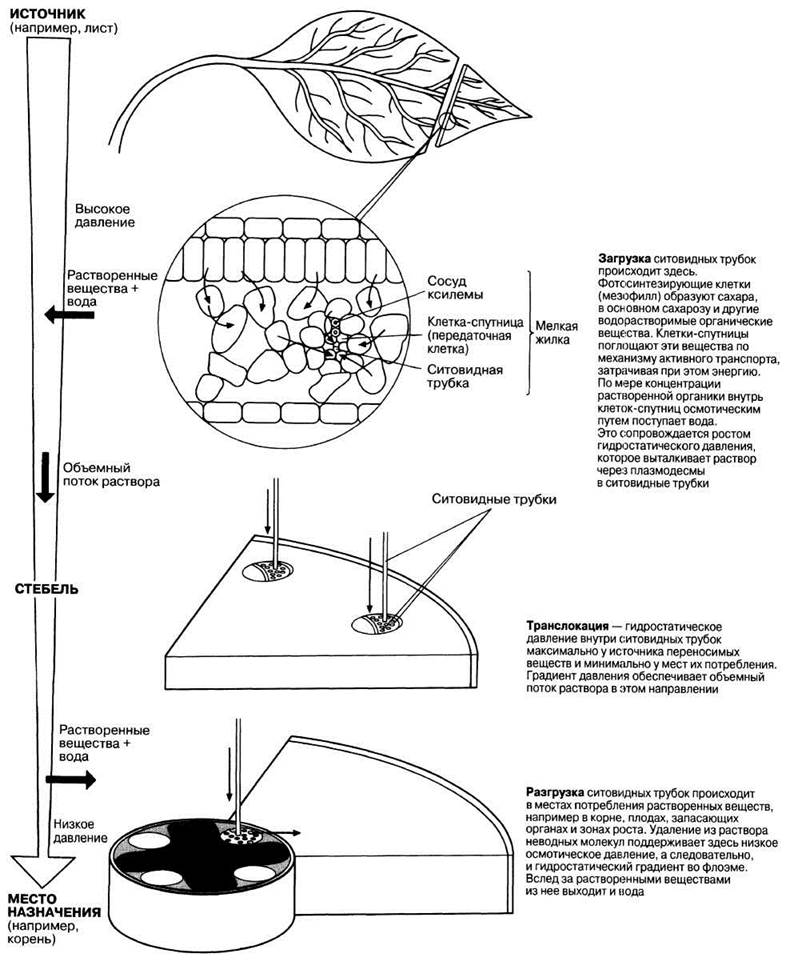
Рис. 13.29. Передвижение растворимых органических веществ, например сахарозы, по флоэме зеленого растения. Такое передвижение осуществляется в три этапа, а именно: движение растворенных веществ от фотосинтезирующих клеток к ситовидным трубкам (их загрузка), транслокация по флоэме и разгрузка в местах, где эти вещества используются.