Біофізика і біомеханіка - В. С. Антонюк - 2012
Розділ 3. МОЛЕКУЛЯРНА БІОФІЗИКА
3.4.Біологічні мембрани
3.4.4.Радіоспектроскопічні методи дослідження біологічних мембран
Розглянемо два найчастіше використовувані в біофізиці і, зокрема, для дослідження властивостей мембран радіочастотні методи - електронний парамагнітний резонанс (ЕПР) та ЯМР.
Електронний парамагнітний резонанс може спостерігатися на таких парамагнітних частинках:
- атомах і магнітних частинках з непарним числом електронів (наприклад, атомах водню, азоту);
- вільних радікалах хімічних сполук з неспареними електронами.
Оскільки молекули фосфоліпідів не парамагнітні (вони діамагнітні),
у ліпідні мембрани під час дослідження їх методом ЕПР уводять спін- зонди - молекули або молекулярні групи з неспареними електронами (парамагнітні центри). ЕПР-спектри поглинання на введених зондах надають інформацію про оточення ліпідів та їх рухливість. Істотним недоліком ЕПР-методу на спін-зондах є те, що внесення до мембрани сторонніх домішок-зондів може змінити структуру і властивості досліджуваного об’єкта.
Ядерний магнітний резонанс спостерігається, коли на речовину, що містить певні ізотопи, діє сильне постійне магнітне поле і перпендикулярне до нього слабке змінне електромагнітне поле. Ядерний магнітний резонанс можна спостерігати на ядрах, магнітний момент яких відмінний від нуля. Найбільш зручними ізотопами для ЯМР-досліджень є ізотопи 1Н та 12С, оскільки і водень, і вуглець входять майже в усі біологічні об’єкти.
Суть цих методів можна описати, ґрунтуючись на класичних та квантово-механічних уявленнях. Зупинимося лише на класичному поясненні цих методів.
У магнітному полі (зовнішньому або внутрішньому) магнітний момент парамагнітної частинки (ЕПР) або ядра (ЯМР) процесу навколо напрямку магнітного поля з частотою ω0 = γН, де γ - гіромагнітне відношення, яке дорівнює відношенню магнітного моменту частинки до її механічного моменту. Це частота власних коливань системи.
Якщо на систему, що містить частинки, накласти зовнішнє змінне поле частотою ω, то в разі збіжності частот ω0 = ω спостерігатиметься резонанс. Резонанс - явище різкого підвищення потужності, що поглинається від радіочастотного джерела, якщо його частота збігається з частотою власних коливань системи.
Для спостереження резонансу необхідно змінювати або частоту зовнішнього джерела змінного поля, або частоту власних коливань системи, ν > 0 (змінюючи величину зовнішнього поля Н).
Спектрами ЕПР та ЯМР називають залежність поглиненої потужності електромагнітного поля Р від величини зовнішнього поля Н. Характерний вид спектра показано на рис. 3.14. Форма ліній спектрів ЯМР та ЕПР зазвичай має вигляд гаусових або лоренцових кривих.
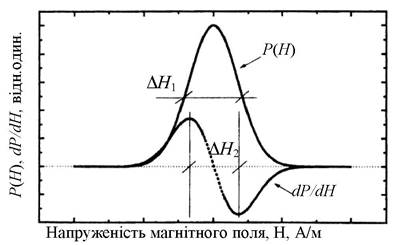
Рис. 3.14. Залежність резонансно поглиненої потужності Р(Н) і похідної від потужності dP/dH від зовнішнього поля Н
Для збільшення чутливості експериментальної установки і точності визначення параметрів системи зазвичай використовують не поглинювану потужність, а похідну від потужності залежно від dP/dH. Для отримання похідної від потужності зовнішнє магнітне поле модулюють за амплітудою. Величина модулювального поля має бути такою, щоб за максимальної величини сигналу не було спотворень форми спектральної лінії або її розширення. Структурну схему ЯМР спектрометра показано на рис. 3.15.
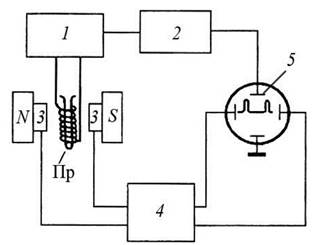
Рис. 3.15. Блок-схема ЯМР спектрометра: 1 — автодонний НВЧ-генератор; 2 — підсилювач; 3 — модуляційні котушки та котушки електромагніту; 4 — блок модуляції; S — індикатор спектрів N-S — полюси постійного магніту; Пр — пробірка зі зразком та намотаною навколо неї котушкою індуктивності
Спектри характеризуються такими параметрами: кількістю і розміщенням резонансних ліній, інтенсивністю ліній, шириною ліній.
Припустімо, що сигнал ЕПР може спостерігатися на декількох парамагнітних частинках, що входять в досліджувану систему. Парамагнітна частинка займає декілька нееквівалентних позицій в системі. Тоді кожній позиції від
повідатиме своя лінія поглинання в спектрі ЕПР, інтенсивність кожної лінії визначатиметься відносною заселеністю відповідної позиції.
Аналогічна картина спостерігається і під час досліджень ЯМР. Так, якщо, наприклад, водень займає декілька нееквівалентних позицій в молекулі зразка, то кожній його позиції відповідатиме лінія в ЯМР-спектрі. Наприклад, ЯМР-спектр на ізотопі 1Н етилового спирту С2Н5ОН складається з трьох ліній, які відповідають групам атомів ОН, СН2 та СН3 з інтенсивностями, що відносяться як 1:2:3, відповідно кількості атомів водню, що входять у ці групи (рис. 3.16).
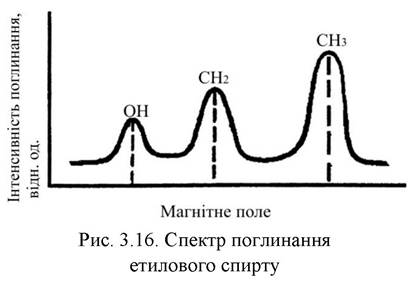
Ширина ліній спектрів ЯМР визначається силою взаємодії між молекулами зразка. Чим сильніша взаємодія, тим ширша лінія. Рух ядер приводить до звуження ліній.
Розглянемо електрон, що обертається по колу радіусом г з частотою V. Тоді механічний момент електрона P = mevr, 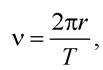 де Т - період обертання електрона на орбіті. Звідси
де Т - період обертання електрона на орбіті. Звідси 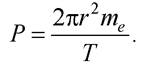
Магнітний момент електрона M = IS2, де I - мікрострум, зумовлений рухом електрона по електронній орбіті навколо ядра, S = πr2 - площа контура, що охоплюється струмом. Величина мікроструму І дорівнює заряду електронів відносно періоду обертання електрона за одиницю часу:
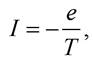
де е - заряд електрона; Т - період обертання електрона по орбіті. Тоді магнітний момент
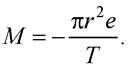
Магнітомеханічне відношення визначають так: γ = gγ0, де g - фактор Ланде; γ0 - одиниця магнітомеханічного відношення; для електрона
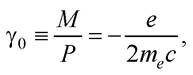
де с - швидкість світла у вакуумі; с = 3 · 108 м/с.
Тоді частота прецесії магнітного моменту можна подати у вигляді
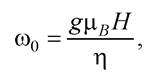
де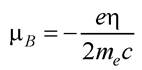 - магнетон Бора, зумовлений рухом електрона;
- магнетон Бора, зумовлений рухом електрона;
μ5 = 9,27 · 10-28 Дж/Тл.
У магнетонах Бора вимірюється магнітний момент системи, зумовлений орбітальним рухом та спіном електронів.
У випадку ЯМР наведені вище міркування аналогічні, а частота прецесії
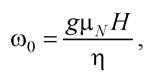
де μN - ядерний магнетон;![]()
Високочастотна електромагнітна хвиля, яка падає на зразок, що вивчається, порушує рівновагу в системі спінів. Релаксація спінів до рівноважного стану характеризується часом релаксації Т1, Т2, від якого залежить ширина лінії спектра ЕПР:
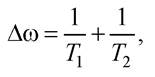
де T1 - час спін-ґраткової релаксації; Т2 - час спін-спінової релаксації. Час T1 визначається швидкістю встановлення рівноваги між системою спінів і ґратками, або взаємодією магнітних моментів частинок з тепловими коливаннями атомів. Величина Т1 залежить від температури, оскільки зі зміною температури змінюються амплітуди коливань атомів. Величина Т2 визначається швидкістю рівноваги в системі спінів і від температури майже не залежить.
Ширина ліній спектрів ЯМР та ЕПР залежить від в’язкості досліджуваної системи. За нормальних умов (кімнатної температури, атмосферного тиску) і нормального фізіологічного стану ліпідна частина мембрани є рідкою, її в’язкість становить 30...100 мПа · с (для порівняння: в’язкість води становить 1 мПа-с). Зміна мікров’язкості ліпідної час
тини біологічних мембран призводить до патологічних змін клітин і є предметом ЯМР та ЕПР досліджень. Зокрема, прикріпляючи спін-зонди до кінців ліпідних хвостів та полярних головок методом ЕПР, було встановлено, що рухливість ліпідних хвостів біля їхніх кінців вища, ніж біля полярної головки.
Рухливість ліпідних молекул досить велика. При цьому розрізняють латеральну дифузію та фліп-флоп дифузію - дифузію поперек мембрани. Латеральну дифузію досліджують під мікроскопом. Швидкість фліп-флоп дифузії визначають методом ЕПР за допомогою спін-зондів на ліпосомах. Фосфоліпідні молекули, що формують ліпосоми, виявляють спін- зондами. Після того в систему додають аскорбінову кислоту. Взаємодія не- спарених електронів з кислотою переводила парамагнітні молекули в діамагнітний стан, що зменшувало інтенсивність сигналу ЕПР. Безпосередньо після дії аскорбінової кислоти інтенсивність ЕПР спектрів зменшувалася в два рази. Це відбувається унаслідок переходу молекул, що перебували на зовнішніх поверхнях ліпосом, з парамагнітного в діамагнітний стан. Надалі інтенсивність лінії ЕПР повільно зменшувалася унаслідок фліп-флоп дифузії мічених молекул з внутрішнього боку мембрани на зовнішню. За час, який становить близько 6,5 год, величина сигналу зменшується в два рази, що й визначає середню швидкість поперечної дифузії.