Біофізика і біомеханіка - В. С. Антонюк - 2012
Розділ 3. МОЛЕКУЛЯРНА БІОФІЗИКА
3.4.Біологічні мембрани
3.4.3. Фізичний стан і фазові переходи ліпідів у мембранах
Суміші ліпідів з водою мають властивості поліморфізму. Це може бути ламелярна фаза гелю, ламелярна рідкокристалічна фаза, гексагональна фаза типу II, кожна з яких буде залежати від концентрації ліпідів, температури, тиску, ліпідного складу, іонної сили та кислотності (рис. 3.12).
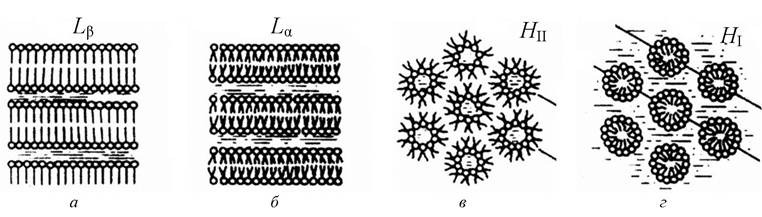
Рис.3.12. Схематичне зображення різних фаз водно-ліпідних систем: а - ламелярна гелева фаза; б - ламелярна рідкокристалічна фаза; в - гексагональна фаза типу II; г - гексагональна фаза типу I
За нормальних фізіологічних умов мембрани перебувають у рідкому стані, проте на відміну від рідин вони мають упорядковану просторову структуру. Завдяки таким властивостям стан мембран називається рідкокристалічним.
1. Ламелярна рідкокристалічна фаза Lα. Для цієї фази характерне впорядковане розміщення шаруватих структур за значної невпорядкованості ацильних ланцюгів молекул. Саме в цій фазі перебуває основна маса ліпідів у біомембранах.
2. Ламелярна гелева фаза Lβ. Зі зниженням температури мембрани переходять з рідкокристалічного стану в твердокристалічний (гель- стан). За такого переходу зберігається загальна структура мембрани, але порядок у системі ще більше зростає. Якщо в рідкому стані площа мембрани, що припадає на одну молекулу ліпіду, становить 0,58 нм , то в гель-стані ця величина зменшується до 0,48 нм . Товщина мембрани у разі переходу в твердокристалічний стан збільшується, але за рахунок зменшення площі об’єм мембрани в цілому зменшується. Молекули упаковані щільніше (на молекулу припадає менша площа поверхні), ацильні ланцюги більш упорядковані і перебувають повністю в транс- конфігурації. Щільність і товщина бішару у фазі гелю більші, ніж у рідкокристалічній фазі (молекули максимально витягнуті).
3. Гексагональна фаза I (Ні). Ліпідні молекули формують циліндрові структури, поверхня яких утворена полярними головками, що контактують з водою. При цьому циліндри паковані з утворенням гексагональних ґраток.
4. Гексагональна фаза II (Н). Ліпіди також утворюють циліндри, але полярні головки ліпідів обернені всередину циліндрів, де міститься вода. Паковання циліндрів також гексагональне.
Фазовий стан, а головне в’язкість ліпідного бішару мембран, впливає на каталітичну активність мембранних ферментів, на проникність мембран, а отже, на процеси обміну речовин у клітинах. Чим більша рухливість молекул фосфоліпідів, тим вища в’язкість мембран і тим краща проникність для дифундувальних речовин. Під час переходу мембранного бішару в стан гелю швидкість латеральної дифузії фосфоліпідів зменшується більш ніж на два порядки (D < 104 см2/с).
Рухливість ліпідних молекул в обох фазових станах істотно різняться. У гель-стані ліпіди здатні здійснювати тільки сумісні коливання або обертальні рухи. В рідкому стані ліпідні хвости мають набагато більшу свободу, особливо велика їх рухливість усередині мембрани.
Під час фазового переходу збільшується проникність мембрани для іонів і низькомолекулярних сполук, здатних проходити крізь такі пори. Фазовий перехід у мембранах відбувається не миттєво, а протягом деякого температурного інтервалу. Температурою фазового переходу називають температуру, за якої одна половина мембранних ліпідів перебуває в рідкокристалічному стані, а друга - у твердокристалічному.
Таким чином, для нормального функціонування мембрана має бути в рідкокристалічному стані. За досить низьких температур ліпідний бішар перебуває в квазікристалічному стані (гель-стані). У разі підвищення температури спостерігається перехід «гель-рідкий кристал». Зміна властивостей ліпідів відбувається у вузькому температурному інтервалі, що характерний для фазового переходу, який спостерігається під час плавлення твердого тіла. Температура фазового переходу знижується з підвищенням ступеня ненасиченості зв’язків між вуглецевими атомами вуглеводневих хвостів ліпідних молекул. Так, температура фазового переходу «гель-рідкий кристал» для мембран з ненасичених ліпідів може становити мінус 20 °С, а для насичених ліпідів - плюс 60 °С. Тому в разі тривалого зниження температури мікроорганізмів, рослинних і тваринних клітин спостерігається адаптаційна зміна хімічного складу мембран для зниження температури фазового переходу. Передбачається, що на первинний механізм кріогенних руйнувань біомембран впливає фазовий перехід у гель-стан. Дослідження ролі холе- стеролу в біомембрані показали, що в рідкокристалічному стані суміші ліпідів холестерол обмежує конформаційну рухливість фосфоліпідних ланцюгів. У стані гелю він затруднює оптимальне паковання ланцюгів у повну трансконфігурацію, знижуючи сили тяжіння між вуглеводневими ланцюгами ліпідів. У результаті суміші фосфоліпід-холестерол за впорядкованістю займають проміжне положення між станами гелів і рідкокристалічних систем чистого фосфоліпіду. Таким чином, наявність холестеролу може зменшувати зміни в мембрані, які супроводжують фазовий перехід.
Фазові переходи типу «гель-рідкий кристал» відбуваються за температури Тф.п (рис. 3.13), величина якої залежить від умісту води в системі.
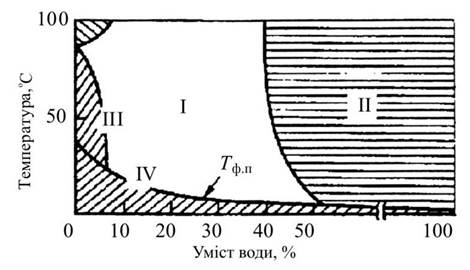
Рис. 3.13. Фазова діаграма суміші яєчного лецитіну II — двофазна система: вода, бішар; III — ділянка
існування гексагональних структур; IV — гель; Тф.п — крива температури фазового переходу
Так, Тф.п досягає мінімуму, коли загальний вміст води перевищує ту кількість, яку можуть зв’язувати ліпіди (концентрація ліпідів мінімальна).
Проте за температури вищої за Тф.п і нестачі води ліпіди можуть перебувати у впорядкованому стані.
Крім переходів типу «гель-рідкий кристал», деякі ліпіди (фосфатидна кислота, фосфотиділсерин) можуть зазнавати перетворення, що приводять до утворення гексагональних структур. Підвищення температури, пониження гідратації бішару, збільшення ненасиченості жирнокислотних ланцюгів, висока іонна сила за лужного рН-чинника, які сприяють утворенню в ньому гексагональних структур. Перехід окремих ділянок у гексагональну фазу призводить до порушення цілісності мембрани, формування каналів проникності тощо.
Живі організми, що мешкають у різних кліматичних умовах, мають різне співвідношення насичених та ненасичених зв’язків у молекулах ліпідів, що забезпечує пристосування його до холоду або жари залежно від середовища їхнього мешкання. Більш того, це співвідношення змінюється для різних ділянок тіла одного й того ж організму. Наприклад [21], температура ноги біля копита полярного оленя може становити мінус 20 °С, а температура ноги біля тулуба досягає +30 °С. Клітинні мембрани не зазнають фазового переходу, оскільки мембрани клітин біля копита містять більше ненасичених ліпідів, а біля тулуба - більше насичених. Відомо, що тваринні жири за кімнатної температури зазвичай перебувають у твердому стані, а рослинні навпаки - в рідкому. Це пояснюється тим, що в тваринних клітинах менше ненасичених ліпідів, ніж у рослинних.
Одним з найпоширеніших методів вивчення фазових переходів у мембранах є метод мікрокалориметрії, який дозволяє визначити кількість теплоти Q, яка була поглинена під час плавлення речовини, що містить ν молів ліпідів. Знаючи Q, можна розрахувати питому ентальпію плавлення: ΔΗ = Q/v.
Таким чином, плавлення речовини відбувається за такої температури Тпл, для якої енергія Гібса в твердому стані (Gтв = Нтв - ТплSтв) дорівнює енергії Гібса в рідкому стані (Gр = Нр - TплSp).