Біофізика і біомеханіка - В. С. Антонюк - 2012
Розділ 3. МОЛЕКУЛЯРНА БІОФІЗИКА
3.4.Біологічні мембрани
3.4.2. Динаміка біологічних мембран
За фізіологічних температур ліпідний бішар мембрани перебуває в рідкокристалічному стані. З одного боку, мембрана має властивості рідини - плинність, лабільність (її окремі компоненти рухомі), а з другого боку, зберігає високий ступінь упорядкованості ліпідних молекул (вони розміщені в два шари і вуглеводневі хвости витягнуті), що характерно для твердих кристалів. У межах кожного шару можливий великий або менший ступінь упорядкованості. Відповідно до дифракційної картини бішар складається з численних кристалітів (добре пакованих ділянок) розмірами близько сотень ангстремів, орієнтація яких дещо відрізняється від орієнтації сусідніх кристалів. Такі утворення мають динамічний характер та видозмінюються з плином часу. Поєднання високої впорядкованості з плинністю і лабільністю забезпечує виконання мембранами їх основних функцій.
Рух ліпідних молекул може бути паралельним та поперечним до поверхні мембрани. Рух, паралельний поверхні, - латеральна дифузія. Це рух у двовимірному просторі бішару, зміна орієнтації полярних головок, коливання жирнокислотних ланцюгів, а також обертальна рухомість навколо довгої осі. Поперечний рух - повільний обмін компонентами моношарів (фліп-флоп), за якого молекула має не тільки переміститися в інше місце, але й повернутися, оскільки її полярна головка повинна бути напрямлена в протилежний бік.
Коефіцієнт латеральної дифузії для ліпідів D = 108...107 см2/с, його значення залежить від складу мембран і температури. З погляду молекулярного руху для інтерпретації експериментальних даних використовують деякі прості моделі руху мембранних компонентів. Поступальна хода ліпідних та білкових молекул подібна до двовимірної дифузії. Ізотропна двовимірна дифузія характеризується середньоквадратичним переміщенням, що відбувається за час t:![]() , де D — коефіцієнт ізотропної дифузії.
, де D — коефіцієнт ізотропної дифузії.
У цьому випадку молекула переміщується за одну секунду на відстань 2 мм у середовищі з дифузією Dліп ≈ 10-8 см2 /с.
Частота перескоків молекул (с-1) унаслідок латеральної дифузії
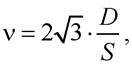 де D - коефіцієнт латеральної дифузії; S - площа однієї молекули на поверхні мембрани. Для молекул ліпідів S = 6 ·10-5 см , тоді ν = 107 с-1.
де D - коефіцієнт латеральної дифузії; S - площа однієї молекули на поверхні мембрани. Для молекул ліпідів S = 6 ·10-5 см , тоді ν = 107 с-1.
Час осілого життя молекули в одному положенні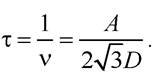
Отже, середній час одного перескоку t =10-7 с. Час обертального руху фосфоліпідів у природних мембранах t = 10-6 с.
Рухливість молекул білків у мембрані трохи менша. Коефіцієнти латеральної дифузії для білків: Dбіл = 10-2 …10-10 см2/с, обертальна рухливість білків характеризується часом tоб = 10-6…10-4 с.
Приклад 3.1. Знайдіть середню величину зміщення молекули формаміду у воді та у розчині сахарози за 1 хв, якщо коефіцієнти дифузії цієї речовини у воді й сахарозі відповідно становлять 1,6·10-5 і 0,3·10-5 см2с-1 .
Розв’язання:
Середньоквадратичне переміщення молекул:
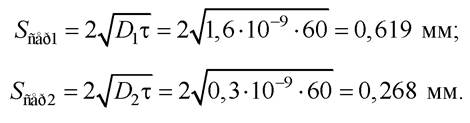
Коефіцієнт дифузії ліпідів дуже великий, при цьому ліпіди, що перебувають поблизу молекул білків у межах двох-трьох діаметрів ліпідної молекули, дифундують з дещо меншою швидкістю, ніж вільні ліпіди. Молекули білків також досить швидко переміщуються, хоча й повільніше, ніж ліпіди.
Порівняно з латеральною дифузією «фліп-флоп» переходи відбуваються набагато рідше. Середній час між переходами молекули фос- фоліпіду з однієї поверхні мембрани на іншу не перевищує однієї години. Молекули білків майже не здійснюють таких переходів.
Істотні відмінності між швидкостями латеральної дифузії і «фліп- флоп» переходів дуже важливі. Така велика швидкість латеральної дифузії забезпечує високу швидкість хімічних реакцій, що відбуваються на мембранах, а мала швидкість «фліп-флоп» переходів сприяє збереженню асиметрії мембран.
Завдяки поєднанню швидкої дифузії в площині мембрани з повільною дифузією упоперек мембрани підтримується асиметрія (анізотропія) розміщення молекул ліпідів та білків, певна орієнтація білків- ферментів упоперек мембрани. Це має важливе значення для напрямленого перенесення речовин.
В організмі хребетних, а також багатьох безхребетних тварин існують базальні мембрани - стінки, навколишні групи клітин або рідше окремі клітини, які часто розміщуються між епітеліальною і сполучною тканинами в стінках кровоносних судин. Вони забезпечують механічні зв’язки між клітинами, відіграють роль фільтра, особливо в стінках кровоносних судин, де регулюють проникнення речовин з крові в тканини. Завдяки ньому в мозок не проникає багато речовин, зокрема цілий ряд лікарських речовин.
Експериментальні дослідження показують [95], що мембрана містить певні типи рухомих структурних дефектів - кінків, які є наслідком конформаційних перебудов вуглеводневих ланцюгів. Унаслідок малої величини (близько 3,5 ккал/моль) енергетичного бар’єра для обертання навколо одиничного С-С-зв’язку жирно- кислотних радикалів, велика ймовірність трансгошізомеризації (гошконформація виникає при повороті навколо С-С-зв’язку на ±120° відносно транс-конформації).
Якщо витягнуту молекулу повернути навколо С-С-зв’язку на кут 120°, а потім навколо іншого С-С-зв’язку, то вона розміститься через одну від початкової, на кут мінус 120°, то вийде конформація, названа простим кінком (рис. 3.11).
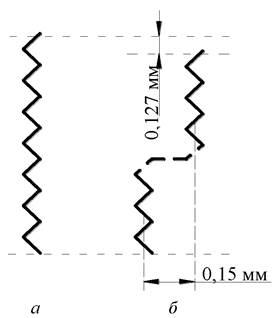
Рис. 3.11. Схематичне зображення вуглеводневих ланцюгів в трансконфігурації (а), трансгошконфігурації (б)
При цьому лінійна молекула виявляється немовби двічі переламаною. Синхронні трансгошперетворення можна подати як рух кінків уздовж ланцюга. Таким чином, наявні у вуглеводневій частині мембрани і кінки утворюють невеликі рухомі «кишеньки» або «бульбашки» вільного простору різного розміру молекул, що транспортуються з водної фази, можуть перейти у порожнину кінка і дифундувати через мембрану разом з рухомим кінком. Наявність у вуглеводневих ланцюгах кінків, подвійних цисзв’язків та інших особливостей визначають рухливість (рідинність або плинність) ліпідного бішару за нормальних умов. При цьому площа ефективного поперечного перерізу ланцюгів збільшується до 0,5 нм (за повної трансконфігурації мінімальне її значення становить 0,19 нм ).