Біофізика і біомеханіка - В. С. Антонюк - 2012
Розділ 3. МОЛЕКУЛЯРНА БІОФІЗИКА
3.5.Транспорт речовини через біологічні мембрани
3.5.2. Поняття активного транспорту речовини
Активний транспорт - це перенесення речовини з місць з меншим значенням електрохімічного потенціалу в місця з його більшим значенням. Активний транспорт у мембрані супроводжується зростанням енергії Гібса, він не може здійснюватися мимовільно, а лише за рахунок витрати енергії, яка запасена в макроергічних зв’язках АТФ.
Активний транспорт речовин через біологічні мембрани має величезне значення. За рахунок активного транспорту в організмі створюються градієнти концентрацій, градієнти електричних потенціалів, градієнти тиску тощо, які підтримують життєві процеси, тобто з погляду термодинаміки активне перенесення утримує організм у нерівноважному стані, підтримує життя.
Активний транспорт через біологічні мембрани досліджував данський учений Г. Уссінг (1949 р.) у дослідах з перенесення іонів натрію через шкіру жаби (рис. 3.19) [56]. Перенесення ґрунтувалося на вимірюванні активного транспорту електричним методом (методом короткого замикання). Якщо розділити дві камери з однаковими розчинами Рінгера шкірою жаби, то між її зовнішньою (мукозною) та внутрішньою (серозною) поверхнями виникає різниця потенціалів, яка приблизно дорівнює 100 мВ (потенціал на зовнішній поверхні буде від’ємним). При цьому спостерігалися потоки іонів натрію крізь шкіру жаби: від зовнішньої до внутрішньої поверхні і від внутрішньої до зовнішньої поверхні. Відношення потоків іонів у разі пасивного транспорту виражається рівнянням Уссінга:
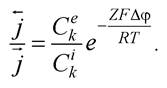
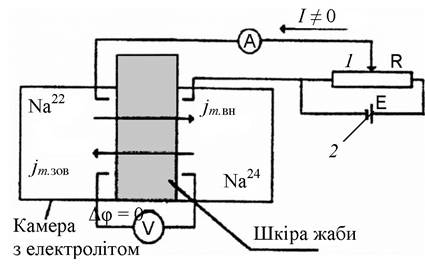
Рис. 3.19. Схема експериментальної установки Уссінга:
А - амперметр, V- вольтметр;
1- реостат; 2- конденсат; 3- гальванічний елемент
Регулюючи струм у компенсувальному колі, можна досягти різниці потенціалів до нуля. Тоді в системі має настати рівновага, оскільки немає градієнтів концентрацій іонів і потенціалу між камерами, розділеними шкірою жаби. В таких умовах пасивне перенесення іонів згідно з критерієм Уссінга неможливе, сумарний потік дорівнює нулю.
Проте під час проведення експериментів було виявлено [94], що струм, який тече в компенсаційному колі, протікає і крізь шкіру. Отже, відбувається одностороннє перенесення іонів. Методом мічених атомів було показано, що потік іонів натрію від зовнішньої до внутрішньої поверхні більший від потоку в протилежному напрямку. Таким чином, струм усередині епітелію створюється активним перенесенням іонів натрію.
Згідно із сучасним уявленням активно транспортуються іонні насоси біологічних мембран - спеціальні білкові комплекси (транспортні АТФ). Відомі три типи електрогенних іонних насосів. Комплекс К+-Nа+-АТФ за рахунок енергії гідролізу однієї молекули АТФ у клітину переносяться два іони калію і виносяться з клітини три іони натрію. Отже, насос генерує електричний струм через мембрану, тому К+-N+-насос - електрогенний. Са+-АТФ переносить два іони кальцію, а Н+-насос - два протони з використанням енергії однієї молекули АТФ.
У більшості моделей активного транспорту передбачається процес сполучення хімічної реакції гідролізу АТФ (скалярного процесу) з напрямленим просторовим переміщенням (векторним процесом). У цьому випадку білок або білковий комплекс повинен виконувати як ферментативну, так і транспортну функції. Отже, у простому випадку він може містити один активний центр і один центр скріплення іонів. Для забезпечення транспорту іонів між двома мембранами передбачається, що центр скріплення іонів може поперемінно контактувати з обома розчинами, розділеними мембраною. Це, наприклад, мінорні рухи, які відкривають та закривають канали, сполучені з розділеними мембраною розчинами, або повернення всієї молекули ферменту в площину мембрани, що зумовлюють пересування центра скріплення іонів через мембрану. Можливі також зміни просторової форми макромолекули для переміщення центра скріплення йонів.
Як показали дослідження, білок K+-Nа+-АТФ існує в двох різних конформаціях — Е1 та Е2, при цьому Е1 має високу спорідненість з Na+ і низьким - з K+ (місця скріплення іонів орієнтовані до цитоплазми). Ця конформація білка має високу спорідненість з молекулами АТФ. Конформація Е2 ферменту має високу спорідненість з К+ і низьку - з Na+ (місця скріплення обернені назовні). При цьому Na+ потрібен для швидкого фосфоритування, а K+ - для швидкого дефосфоритування.
Перехід іонів K+ із зв’язаного з ферментом стану у вільний стан відбувається на завершальному п’ятому етапі. Цей процес індукується скріпленням АТФ з алостеричним центром ферменту, який має низьку спорідненість.
Таким чином, у процесі активного транспорту молекула АТФ виконує дві функції - субстрати та алостеричного ефектора.
Запропоновані моделі будови, згідно з якими ці білкові комплекси містять 6, 8 або 10 трансмембранних α-спіральних сегментів. Є велика гідрофільна ділянка, що містить домени, де зв’язуються нуклеотиди і де відбувається фосфоритування. При цьому зміна конформації, що супроводжується гідролізом АТФ, зумовлює зміну спорідненості відповідних центрів білка з іонами.
Створені за допомогою систем активного транспорту градієнти концентрацій речовин утворюють на мембрані різницю хімічного або електрохімічного потенціалу, за рахунок чого можуть переноситися інші речовини проти градієнтів їх концентрацій. Такий транспорт, джерелом енергії якого є не безпосередньо АТФ або енергія окиснювально-відновних реакцій, а градієнт концентрацій інших речовин називають вторинно-активним, або зв’язаним.
Є три види вторинно-активного транспорту іонів: уніпорт, симпорт і антипорт.
У разі уніпорту за рахунок існування на мембрані градієнта електричного потенціалу здійснюється однонапрямлений транспорт заряджених частинок у бік меншого значення потенціалу. У процесі антипорту (обмінного транспорту) здійснюється транспорт однаково заряджених іонів двох типів у різних напрямках. Згідно з механізмом симпорту (ко- транспорту) здійснюється транспорт протилежно заряджених іонів в одному напрямі. При цьому один з іонів транспортується за градієнтом концентрацій, а другий - за градієнтом електричного потенціалу, який створюється транспортом першого іона.
Одним з найбільш вивчених випадків вторинно-активного транспорту незаряджених молекул є всмоктування глюкози в стінки кишечнику. Тут транспорт глюкози здійснюється проти градієнта концент
рації, але не за рахунок енергії АТФ, а завдяки градієнту концентрації іншої речовини, у цьому випадку - іонів натрію. Подібні системи транспорту існують і для багатьох інших речовин, наприклад вуглеводів і амінокислот, що дуже важливо, оскільки для них немає специфічних насосів.