Біофізика і біомеханіка - В. С. Антонюк - 2012
Розділ 4. БІОФІЗИКА СКЛАДНИХ БІОЛОГІЧНИХ СИСТЕМ
4.1.Математичне моделювання складних біологічних систем
4.1.1.Модель збільшення чисельності популяції
У математичному моделюванні часто використовують диференціальні рівняння, наприклад, коли процеси, що вивчаються, або явища зручно описувати плоскими функціями, коли в описі беруть участь такі поняття, як швидкість, пришвидшення. Тут мають на увазі не швидкість якої-небудь точки або тіла, а швидкість перебігу процесу: це можуть бути процеси наповнення, поширення тощо. Звичайні диференціальні рівняння поширені в інженерній практиці, кожне з них може описувати який-небудь інженерний процес.
Розглянемо дві математичні моделі, які дозволяють знайти залежність зміни чисельності популяції від часу для різних умов функціонування системи.
Модель природного зростання чисельності популяції (модель Мальтуса). Реальна система: є деяка популяція одного вигляду (мікроорганізми, зайці тощо), у якій відбуваються життєві процеси у всьому їх різноманітті.
Постановка завдання. Знайти закони зміни чисельності популяції в часі.
Основні припущення:
1) існують тільки процеси розмноження і природної загибелі, швидкості яких пропорційні чисельності особин у певний момент часу;
2) не враховуються біохімічні, фізіологічні процеси;
3) не відбувається боротьба між особинами за житло, за їжу (нескінченно великий простір та кількість їжі);
4) розглядається тільки одна популяція, немає хижаків.
Модель. Величини: х - чисельність популяції у момент часу t; R - швидкість розмноження; γ - коефіцієнт розмноження; S - швидкість природної загибелі; σ - коефіцієнт природної загибелі; dx/dt - швидкість зміни чисельності популяції; ε - коефіцієнт зростання.
Тоді R = γx, S = -σx.
Зміна чисельності особин за одиницю часу визначається кількістю народжених за цей час особин та особин померлих: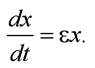
Початкова умова: якщо t = 0, чисельність особин х = х0: х = x0eεt.
Аналіз розв ’язання:
а) ε < 0 (якщо σ > γ), тобто швидкість загибелі більша за швидкість розмноження. Чисельність особин з плином часу зменшиться до нуля;
б) ε > 0 (якщо γ > σ), тобто швидкість розмноження більша за швид
кість загибелі. Чисельність особин необмежено збільшується з плином часу;
в) ε = 0 (якщо σ = γ), тобто швидкість загибелі дорівнює швидкості розмноження. Чисельність особин не змінюється, залишаючись на початковому рівні.
Модель, якщо ε > 0, адекватна реальності лише до певних значень часу. Згідно з цією моделлю, за якою розглядається зменшення чисельності особин тільки за рахунок природної загибелі, їх чисельність має нескінченно збільшуватися з плином часу, що не відповідає реальності.
Модель зміни чисельності популяції з урахуванням конкуренції між особинами (модель Ферх’юльста). Нехай відбувається боротьба між особинами, наприклад, за житло, тим самим додається додаткове джерело загибелі. Вважаючи, що швидкість загибелі за рахунок конкуренції між особинами пропорційна ймовірності зустрічей двох особин, можна записати S = -5 хх - σx (δ - коефіцієнт пропорційності). Тоді рівняння балансу чисельності особин: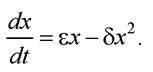
Це нелінійне диференціальне рівняння. Замінивши змінні:
![]()
і врахувавши, що за значення t = 0 величина х = х0, отримують:
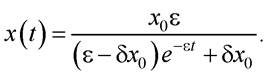
Видно, що з плином часу значення х не прямує до нескінченності, а досягає стаціонарного рівня: хст = ε / δ.
Обидві наведені моделі є основою моделювання процесів у біотехнології (наприклад, для встановлення оптимальних режимів вирощування різних мікроорганізмів).
Розглянемо деякі положення теорії епідемій. Нехай захворювання має тривалий характер, а процес передавання інфекції значно швидший, ніж тривалість хвороби. Розглядатимемо процес передачі інфекції, при пускаючи, що хворі залишаються серед здорових і передають інфекцію при контактах. Припустімо, що m і n - відповідно кількість заражених та незаражених у початковий момент часу; x(t) - кількість незаражених у момент часу t, а y(t) - кількість заражених до цього ж моменту часу. Для всіх моментів часу з деякого досліджуваного проміжку часу, який менший за час життя одного покоління, справджується очевидна рівність: x - y = m + n. Оскільки інфекція передається при контактах, тоді кількість незаражених з часом зменшуватиметься пропорційно кількості зустрічей (наприклад, у разі розповсюдження СНІДу), тобто пропорційно добутку ху. Звідси можна вивести диференціальне рівняння:

де k - коефіцієнт пропорційності. Підставивши в рівняння (4.1) у, виражене через х, отримують диференціальне рівняння:
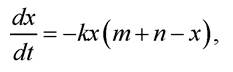
яке має розв’язок такого вигляду:
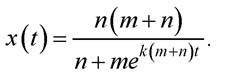
Цей розв’язок визначає закон спадання x(t) з плином часу.
Завдання ускладниться, якщо припустити, що хвороба нетривала і ті, хто видужав, набувають імунітету. Потрібно буде розв’язувати систему диференціальних рівнянь. Ще більше ускладниться завдання, якщо припустити перенесення інфекції паразитами-переносниками (наприклад, малярії - комарами). Кількість рівнянь у системі збільшується. В цьому разі доведеться використовувати ПЕОМ.
Як приклад можна навести зростання дерева і розглянути його як вільну енергії, яка створює фотосинтез у зеленій частині рослин. Ця енергія буде пропорційна площі поверхні зеленої частини рослини (листя) і її витрату можна подати так:
- частина енергії - на підтримання самого процесу фотосинтезу; вона пропорційна площі зеленого покриву, але з іншим (меншим) коефіцієнтом пропорційності;
- частина енергії - на транспортування живильного розчину по всій рослині, ця витрата залежить від кількості шляхів транспортування, що визначається здебільшого об’ємом рослини; крім того, частина енергії витрачається на подолання сили тяжіння і зростає зі збільшенням висоти підіймання живильної речовини;
- частина енергії, що залишилася, - на збільшення маси рослини (тобто на саме зростання), вона пропорційна швидкості зростання. Виходячи з відомого закону збереження енергії за певних припущень складають баланс енергії - витрату енергії прирівнюють до її припливу. Розв’язавши отримане диференціальне рівняння, отримують закон зростання дерева (крива на рис. 4.2).
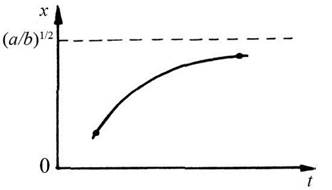
Рис. 4 2 Закон зростання дерева
Із плином часу крива зростання наближається до деякої величини, яка визначає граничну висоту дерева певної породи. Відкладені по вертикальній осі
величини![]() Є деякими кількісними характеристиками породи дерева. Наприклад, знаючи середнє значення висоти дерева певної породи за два вікові періоди (20 та 50 років), можна з розв’язання згаданого диференціального рівняння знайти а і b і побудувати криву зростання. Ця теорія добре узгоджується з експериментальними даними [51].
Є деякими кількісними характеристиками породи дерева. Наприклад, знаючи середнє значення висоти дерева певної породи за два вікові періоди (20 та 50 років), можна з розв’язання згаданого диференціального рівняння знайти а і b і побудувати криву зростання. Ця теорія добре узгоджується з експериментальними даними [51].