АДАПТАЦІЯ РОСЛИН ДО АНТРОПОГЕННИХ ЧИННИКІВ - 2017
4. ДІЯ ІОНІЗУЮЧИХ ВИПРОМІНЮВАНЬ НА РОСЛИННІ ОРГАНІЗМИ
4.3. Загальні закономірності радіобіологічних реакцій рослинного організму
У розмаїтості радіобіологічних реакцій рослини на опромінення є і загальні риси ефекту, які дозволяють убачати наявність багатокомпонентного радіаційного синдрому (Рис. 13) і виділяти такі його вираження:
1. Реакції рослини, які полягають в посиленні ростових і формотворних процесів, що спостерігаються, як правило, за дії малих доз радіації. Ця реакція, названа радіостимуляцією, виявляється або як минуща, короткострокова, або як тривала, що захоплює значну частину вегетаційного періоду. Реакція радіо- стимуляції відрізняється гармонічною інтенсифікацією усіх фізіологічних і пов’язаних з ними біохімічних процесів.
2. За більш високих доз опромінення у рослин розвиваються порушення морфогенезу, обумовлені інактивацією меристем. Ці порушення носять характер морфологічних аномалій, радіоморфозів, а також порушення звичайного ходу формоутворення, відмирання коренів та пагонів, розтягнення вегетаційного періоду, відсутності репродуктивної фази розвитку. Загибель рослинного організму у відповідь на опромінення настає за дуже високих доз радіації і спостерігається не відразу після опромінення, але через певний проміжок часу, протягом якого гинуть меристематичні тканини.
3. Біохімічні і фізіологічні реакції на опромінення виявляються зміною напруженості багатьох процесів і порушенням їхньої супряженості, у результаті чого має місце аномальне нагромадження багатьох продуктів проміжного метаболізму. На порушення біохімічних процесів впливають як інактивація окремих ферментних систем, так і ураження загальної регуляції процесів в опроміненій рослині.
4. Характерною рисою радіаційного синдрому у рослин є утворення генетичних ушкоджень, які виявляються у формі соматичних мутацій, генних мутацій, хромосомних аберацій, геномних мутацій. Вони реалізуються у наступних поколіннях. Порушення хромосом служать причиною інтенсивного клітинного добору у меристемах.
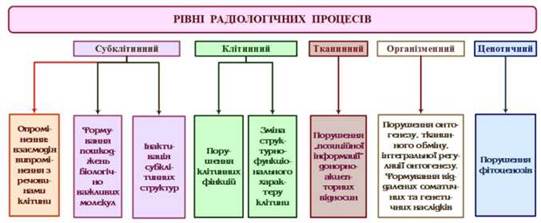
Рис. 13 - Рівні дії радіаційного випромінення за Д. М. Гродзинським
У радіаційному синдромі рослини можна бачити риси спільності із синдромом тваринного організму: наявність критичних тканин і органів, однакові типи цитогенетичних ушкоджень, втрата контролю над інтегральними функціями організму, утворення соматичних мутацій, трансформація клітин і радіаційний канцерогенез.
Очевидно, спільність рис радіобіологічного синдрому відбиває тотожність первинних процесів взаємодії іонізуючих випромінювань з речовинами живих клітин, подібність основних клітинних радіобіологічних реакцій, проліферативної загибелі стовбурних клітин. Розходження виникають там, де зникає подібність процесів рослинних і тваринних організмів.
Звичайно, біологічна реакція на опромінення спостерігається на більш пізніх етапах формування радіобіологічного ефекту, коли процеси його формування захопили рівень клітинних і тканинних явищ. Настання такої реакції спостерігається через різні проміжки часу після дії радіації, у зв’язку з чим говорять про близький і віддалений радіобіологічні ефекти (Рис. 14). Близькі ефекти виявляються безпосередньо в клітинах, підданих опроміненню. Віддалені можуть проявитися в ряді клітинних поколінь від опромінених клітин. Очевидно, віддалені ефекти спостерігаються в тому випадку, коли дози опромінення не перевищують того рівня, після якого всі меристемні клітини цілком втрачають здатність до ділення. Можна і по-іншому визначити сутність близьких і віддалених ефектів. Використовуючи тільки часові характеристики процесів, можна вважати близькими реакції, які відбуваються негайно за гострим опроміненням і, отже, обумовлені радіаційно-хімічними процесами, а віддаленими - реакції, які вимагають для свого прояву деякого лаг-періоду. Наприклад, близьким ефектом є зміна проникності мембран рослинної клітини, яка спостерігається відразу після за опроміненням, віддаленим - утворення мікроядер у клітинах або розвиток морфологічної аномалії.
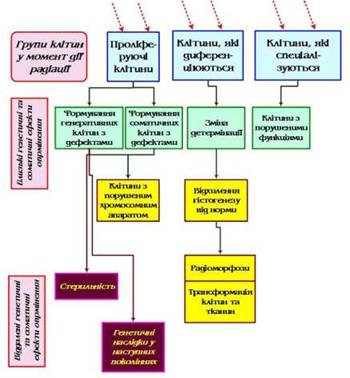
Рис. 14 - Радіаційні ефекти за Д. М. Гродзинським
Генетичні ефекти - генні та соматичні мутації, обумовлені радіаційним ушкодженням клітин, у поколіннях яких виникають або генеративні клітини - мікро- і макроспори - у першому випадку, або ділянка тканини - у другому. Для цього необхідно, щоб здійснилося кілька поділів опромінених клітин, якщо мова йде про вищу рослину. Отже, такі клітини повинні якийсь час нести «сховане» ушкодження, яке згодом реалізується у формі віддаленого ефекту - стерильності пилка або яйцеклітини, появою генної мутації і т. п. Тому з віддаленими ефектами асоціюється трохи формалізоване поняття «схованого» ушкодження. Утім, у цьому понятті немає нічого загадкового: за належного вибору методу завжди можна виявити порушення молекулярних структур, які обумовлюють збереження «схованого» ушкодження.
У складних меристемах вищих рослин утворення більшої частини хромосомних аберацій супроводжується виключенням цих клітин з групи проліферуючих. У цьому механізмі виявляється один зі способів відновлення рослини після променевого ураження.
Порушення процесу детермінації клітин, процесу вибору шляхів диференціації у спеціалізовані тканини може наставати не відразу після опромінення, а після кількох клітинних поділів, які завершаться настанням віддалених соматичних наслідків опромінення - виникненням радіоморфозів, припиненням росту, порушенням формоутворення. У випадку зміни позиційної інформації під час опромінення «сховане» ушкодження полягає у зміні гормонального статусу та порушенні компетентності клітин, тобто має фізіологічний характер. Повертаючись до уявлень про наявність лаг-періоду в радіобіологічних реакціях рослини, можна сказати, що у випадку віддалених реакцій на опромінення лаг-період відповідає інтервалові часу, протягом якого зберігається «сховане» ушкодження. Ще один приклад віддаленого наслідку - трансформація клітин, що може наставати після багатьох клітинних поділів зовні нормальної клітини, що несе «сховане» ушкодження, у якийсь момент спонукаючих клітину здійснювати трансформацію. Звичайно, розкриття природи «схованих» ушкоджень складає дуже важливу проблему сучасної радіобіології.
Віддалені соматичні ефекти - морфози, відсутність репродуктивної фази розвитку за своєю природою близькі до генетичних ефектів, оскільки вони визначають цитогенетичні ушкодження клітин, порушення регуляторних систем, які діють на рівні міжклітинних взаємодій. Розходження між генетичними і соматичними радіобіологічними ефектами полягають у тому, що перші відносяться до клітинних ліній, які ведуть до утворення гамет, а другі - утворювальні клітини, нащадки яких складають тканини тієї ж рослини.
Виникнення генних мутацій відносяться до досить віддалених ефектів опромінення. За опромінення насіння змінені форми виявляються у першому поколінні. Очевидно, радіаційна мутація виникає в ініціалі зародка насіння. «Сховане» ушкодження проходить складний багатоетапний шлях через меристему чекання до спорогенної тканини, далі - до гамети, запліднення і, нарешті, реалізується в новому зародку. Радіація як мутагенний фактор у своєму прояві схожа з хімічними мутагенами. У зв’язку з цим у багатьох роботах радіаційний і хімічний мутагенез розглядаються як явища одного порядку.
Близькі і віддалені ефекти рослини, індуковані іонізуючою радіацією, більш-менш зрозумілі у випадку гострого опромінення організму. За хронічного опромінення картина значно ускладнюється, і для виділення близьких і віддалених ефектів доводиться звертатися до дослідів з гострим опроміненням, що показують, які ефекти подібні за гострого та хронічного опромінення, відносяться до близьких і віддалених. Однак для хронічного опромінення відомі дії, властиві тільки йому. У цьому випадку віддаленими ефектами є ті реакції на опромінення, що настають після певного лаг-періоду. У формуванні віддалених генетичних і соматичних ефектів виявляється безсумнівний вплив відбудовних процесів. Процеси пострадіаційного відновлення полягають у ліквідації наслідків опромінення на певних етапах складного ланцюга подій, що приводять до прояву радіобіологічного ефекту. У ході відновлення ці ланцюги можуть перериватися. Пострадіаційне відновлення може поширюватися на «сховані» ушкодження, що позначається зниженням ступеня прояву віддаленого ураження. У випадку близьких реакцій на опромінення пострадіаційне відновлення також знижує їхню реалізацію.
Закон Бергоньє-Трибондо. Під час зіставлення рівнів радіостійкості різних клітин легко знайти, що для даного виду організмів найбільшою радіочутливістю відрізняються клітини, які знаходяться в стані активної проліферації, що супроводжується високою напруженістю метаболічних процесів. Дійсно, клітини спочиваючих бруньок незрівнянно більш радіостійкі, ніж клітини активних апексів. Зародки сухого насіння більш радіостійкі, ніж набряклого, яке почало проростання. Звичайно, ця закономірність виявляється тільки під час порівняння клітин одного виду рослини. Наприклад, спочиваючі клітини зародків у насінні бобів, як правило, менш радіостійкі, чим активно проліферуючі клітини протонеми мохів. Однак для даного організму незмінно дотримується правило: з підвищенням рівня клітинної активності радіочутливість клітин зростає. Ця закономірність була встановлена ще на початку розвитку радіобіології. Її сформулювали французькі вчені Бергоньє і Трибондо, і це формулювання увійшло в радіобіологію як закон Бергоньє-Трибондо. В наш час цей закон наповнився новим змістом: підвищення радіочутливості у клітин, які знаходяться в активному стані, обумовлене більш уразливою структурою хроматину.
Клітинне ядро і радіочутливість рослини. Широкі межі коливань радіостійкості живих організмів послужили приводом для пошуків кореляції між рівнем радіостійкості і тими або іншими кількісними характеристиками клітини. З таких характеристик основна увага була приділена параметрам клітинного ядра - його об’єму, розмірам хромосом, вмісту ДНК у ядрі та у хромосомах, числу хромосом, співвідношенню між різними нуклеотидами у ядерній ДНК. Пошуки таких кореляцій беруть початок від теорії мішені, що виходить з уявлень про наявність у клітинах мішеней, які уражаються, і на роль яких претендують насамперед клітинні ядра та їхні молекулярні структури. Уже початкові прості якісні зіставлення радіостійкості клітин і розмірів їхніх ядер указували на те, що клітини з більш великими ядрами і хромосомами звичайно відрізняються підвищеною радіочутливістю. Дійсно, вдається реєструвати чіткий зв’язок між деякими кількісними параметрами клітинного ядра і рівнем радіостійкості рослини. Це ж стосується і організмів інших царств живої природи.
Були згруповані різні види організмів, виділені 4 групи, причому кожна група характеризується прямою лінією регресії, що зв’язує обернено пропорційною залежністю рівень радіостійкості із вмістом нуклеотидів у ДНК клітинного ядра (у геномі). Групи включають наступні організми. Перша група: РНК-віруси та одноланцюгові ДНК-віруси; друга - дволанцюгові ДНК-віруси; третя - організми з гаплоїдними клітинами і четверта група - організми, у яких клітини диплоїдні. Чим складніша структура геному клітини, тим вища її радіочутливість.