АДАПТАЦІЯ РОСЛИН ДО АНТРОПОГЕННИХ ЧИННИКІВ - 2017
4. ДІЯ ІОНІЗУЮЧИХ ВИПРОМІНЮВАНЬ НА РОСЛИННІ ОРГАНІЗМИ
4.4. Складові радіаційного синдрому у рослин
Важко виявити який-небудь процес або морфологічну структуру, яка тією чи іншою мірою не змінювалася б після опромінення у досить високій дозі. Навіть швидкий перелік реакцій вищої рослини на опромінення показує, наскільки багато сторін життєдіяльності рослинного організму піддаються змінам. Хромосомні і хроматидні аберації, пікноз ядра, порушення структури мітохондрій та хлоропластів, зміни проникності мембран - усе це типові порушення клітинних структур, які супроводжуються порушенням функціонального стану клітин.
На рівні органів і цілої рослини виявляється зміна архітектоніки. У коренів може підсилитися розгалуження, обумовлене гальмуванням росту осьової частини, збільшенням активності вторинних меристем, формуванням коренів, які утворюються з клітин перициклу. Порушується утворення кореневих волосків. На будові стебла відбиваються обумовлені радіацією гальмування апікальних меристем та активація сплячих бруньок. Під впливом опромінення можуть змінюватися порядок листорозташування, філотаксису, виникати фасціації, пухлиноподібні утворення. Іноді змінюється тип галуження. Радіобіологічні процеси ведуть до виникнення хибних форм будови листової пластинки, зрощенню декількох листових зачатків, зміни типу жилкування. Іноді листя скручується внаслідок порушення росту пластинки, через це ж виникає зморшкувате листя. У формуванні репродуктивних органів також відзначаються відхилення, які ведуть до утворення каліцтв.
Опромінення позначається на багатьох фізіологічних процесах: прискорюється або гальмується хід онтогенезу, що виявляється за змінами темпів формоутворення. За сильного ушкодження меристем зародка насінини з’являються позбавлені утворювальних тканин проростки, які одержали назву «у-паростків». Змінюється під час опромінення і вміст різноманітних речовин у рослині - фосфорних ефірів цукрів, органічних кислот, амінокислот, нуклеотидів, більш складних сполук - пігментів, фітогормонів, речовин вторинного походження. Усе це призводить до залежної від дози загальної зміни метаболізму - дихання, фотосинтезу, біосинтезу багатьох сполук і біогенезу структур.
В усіх цих реакціях опромінених рослин можна вбачати прояв променевого синдрому як сполучення ознак хвороби, променевого патогенезу.
У широкому наборі ознак променевого ураження рослини можна доглянути обмежене коло причин. В основі ураження рослин лежить загибель клітин утворювальних тканин (меристем). Це призводить до того, що ушкоджується велика частина органів і тканин, тому що всі органи рослинного організму виникають з меристем. Тому повна картина радіаційного синдрому у рослин охоплює широку гаму змін, яка набуває зростаючу розмаїтість, коли опромінення здійснюють у моменти найбільшої активності меристем.
Слід зазначити, що радіаційний синдром у рослин істотно залежить від умов опромінення та стану рослин у моменти дії радіації (Рис. 15).
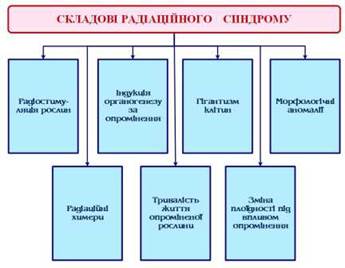
Рис. 15 - Складові радіаційного синдрому
Індукція органогенезу за опромінення. Під впливом радіації часто спостерігаються зміни ходу органогенезу у рослин. Під час опромінення вегетуючої рослини органогенез порушується внаслідок інактивації меристемних клітин.
Фактори, які виникають в опромінених калюсах, можуть переноситися шляхом дифузії через живильне середовище до неопромінених експлантів, індукуючи в останніх органогенез. Отже, логічно припускати, що в індукції органогенезу беруть участь або продукти радіаційно-хімічних реакцій речовин, які утримуються у тканині, або опромінені клітини калюсу продукують речовини типу цитокінінів. Були випробувані індолилоцтова кислота, кінетин, міоінозит, опромінені γ-радіацією у дозах до 250 Гр, у відношенні їхнього впливу на органогенез і виявилося, що всі перераховані опромінені речовини, особливо міоінозит, викликали органогенез у калюсу тютюну. Раніше показано, що опромінена сахароза також набуває здатність продуктами свого радіаційно-хімічного перетворення індукувати органогенез.
У радіаційній індукції органогенезу у рослин варто виділяти як непряму, так і пряму дію випромінювань та ефекти опосередкованого дистанційного впливу випромінювань за допомогою регуляторного впливу, обумовленого зняттям апікального домінування та участю фізіологічно активних речовин, які виникають у результаті дії радіації на тканині.
Гігантизм клітин. За опромінення рослин спостерігають появу аномально великих, гігантських клітин. Такі клітини були виявлені у культурі клітин арахісу (Arachis hypogaea). За опромінення цієї культури дозою 500 Гр цілком придушувалося ділення клітин, але ріст клітин розтяганням продовжувався, у результаті чого до 50-60 % клітин доростали до величезних розмірів. Гігантські клітини характеризуються щільною цитоплазмою, збільшеним числом крохмальних зерен і активним циклозисом. Очевидно, утворення гігантських клітин відбиває втрату контролю над ростом розтяганням. Звичайно, клітини здатні до поділу, починають цей процес, коли досягають визначених розмірів. За опромінення, коли робиться неможливим клітинний поділ, знімається обмеження на граничні розміри клітини.
Явище гігантизму клітин, індукованого опроміненням, вивчено також на водорості едогонії (Oedogonium cardiacum). Гігантські клітини у цієї водорості виникають не тільки безпосередньо з опромінених клітин, але і з дочірніх, які утворюються в результаті поділу опромінених клітин, у яких не виявлялося яких-небудь аномалій поділу. Гігантизм клітин сполучений із блокуванням мітозу. Під час вивчення ультраструктури гігантських клітин водорості виявлені порушення упакування білкових молекул в ультраструктурах клітин, що, можливо, має відношення до контролю морфогенезу. Не виключено також, що за дії іонізуючої радіації ушкоджується цитоскелет, що спричиняє зміну розмірів і форми клітини.
Морфологічні аномалії. Після опромінення насіння рослин у паростків часто спостерігається розвиток морфологічних аномалій - ушкодження окремих органів - листя, стебел. Ці аномалії найчастіше торкаються перших листочків, примордії яких вже розвинулися у зародку насіння і піддалися ураженню. Природа цього явища досить проста: частина меристематичних клітин втрачає здатність до ділення, у результаті чого вони не генерують клітинні потоки, що перешкоджає нормальному формуванню листової пластинки через виникнення стяжок у місцях, де зупинилася генерація нових клітин. Наступне листя нових порядків закладення вже не несуть ознак променевого ураження.
За опромінення вегетуючих рослин морфологічні аномалії можуть виникати у всіх органах, які у момент опромінення перебували у стадії примордиальних бугорків. Усі ці аномалії можна розглядати як меристемогенні. Значно рідше виникають зміни як результат індукованих опроміненням соматичних мутацій локусів, які контролюють морфогенез. Якщо клітини, які мають такі мутації, не елімінують у ході клітинних ділень, то можуть виникати органи зміненої морфологічної структури. Зміни даного типу за своєю природою відносяться до генетичних. Вони можуть мати спадкоємний або морфозний характер.
Радіаційні химери. Унаслідок статистичного характеру розподілу поглиненої енергії у клітинах зародка насіння під час опромінення насіння, паростків або вегетуючих рослин ураження геному різних клітин виявляється неоднаковим. Інакше кажучи, опромінення сприяє тому, що клітини утворювальних тканин, у тому числі й ініціалі, являють собою сукупність ушкоджених клітин з різними дефектами генетичного характеру. Якщо ці ушкодження такі, що поділ клітини не може відбутися, то в ході гістогенезу клітини - носії цих ушкоджень не генерують клітинні лінії і не утворюють відповідні сектори в тканині або органі. Якщо ж індуковані радіацією зміни геному не виключають поділу клітин, то покоління таких клітин входять до складу органів, які формуються, тканин відповідно репродукційної функції тієї або іншої клітини. У результаті рослина, яка виросла з опроміненого насіння, або пагін, який розвився з опроміненої бруньки, складається з генетично різних тканин, являючи собою химеру. У зародку насіння клітини потенційно здатні формувати різні тканини рослини. Оскільки ушкодження, які виникають у ході опромінення, носять статистичний характер, рослини з опромінених однієї і тією же дозою іонізуючої радіації насінин можуть виявитися різними за своєю природою химерами. Для виникнення химери необхідно, щоб соматичні мутації виникали в ініціалі зародка.
Тривалість життя опроміненої рослини. В опромінених рослин може змінюватися тривалість вегетаційного періоду, хоча, як відомо, регуляторні системи, що контролюють темпи проходження фаз розвитку, відрізняються великим консерватизмом і залежать переважно від факторів, сприйняття яких відповідальне за відлік «біологічного часу». Однак, крім орієнтації на хід зовнішніх геофізичних факторів розвиток рослин підкоряється й ендогенному відлікові часу. У результаті накладення сигналів, які надходять ззовні, і внутрішнього контролю темпів розвитку здійснюється, як правило, строго дотримуване часове розгорнення онтогенезу. Опромінення позначається на тривалості вегетаційного періоду, подовжуючи його за підвищених доз. Одну з причин уповільненого розвитку рослин після опромінення можна вбачати в розтягненні тривалості клітинних циклів. Якщо розвиток рослини як послідовний ряд морфогенетичних подій вимагає певного числа клітин, які генеруються апікальними меристемами, то, очевидно, уповільнення клітинних ділень повинно супроводжуватися уповільненням розвитку рослини. У дослідах з опроміненням насіння це помітно по відставанню розвитку паростків з опроміненого насіння порівняно зі звичайними. За дуже високих доз опромінення хід розвитку загальмовується і вегетаційний період розтягується, а іноді рослини не можуть переходити до генеративної фази розвитку. Реакція рослин на фотоперіодичні впливи відрізняється дуже високою радіостійкістю.
Зміна плоїдності під впливом опромінення. Під впливом іонізуючих випромінювань може змінюватися плоїдність клітин рослин. Виникнення тетраплоїдних клітин спостерігали під час опромінення бруньок рослин, які розмножуються вегетативно. Геномна мутація - подвоєння хромосомного набору - не перешкоджає проліферації клітин, і може виникнути пагін, у якому частина клітин (іноді дуже значна) є тетраплоїдними. Частота геномних мутацій буває більше частоти виникнення мутацій інших типів. Так, у винограду (Vitis rotimdifolia) були отримані тетраплоїди, у яких усі клітини внутрішніх тканин були тетраплоїдними, у той час як епідерміс складався з диплоїдних клітин. У шовковичного дерева (Moras nigra) за гострого опромінення пагонів γ-радіацією з’являлися тетраплоїдні клітини, які у ході росту пагону утворювали свої клітинні лінії, у результаті чого виникала химера, що складається з диплоїдних та тетраплоїдних клітин. Радіаційну індукцію тетраплоїдних клітин з диплоїдних виявляли також і в інших рослин.
Під впливом радіації можуть розвиватися і гаплоїдні рослини. Це явище спостерігається в тому випадку, коли квітки запилюють убитим опроміненням пилком, що індукує партеногенез як результат псевдозапліднення. Одержання гаплоїдних рослин зазначеним способом має практичне значення в селекції.
Радіостимуляція рослин. Якщо вивчати дозову залежність ростової функції рослини, вимірюючи її висоту або біомасу, і випробувати вплив зростаючої радіації від найменших, близьких до сотих часток грея, доз, то для багатьох видів рослин виявляється інтервал доз, за яких опромінені рослини відрізняються більш інтенсивним ростом, ніж неопромінені, контрольні рослини. Під час проведення таких експериментів варто досить докладно вивчати інтервал доз від часток грея до декількох десятків грей. Посилення ростової функції рослини за малих доз опромінення одержало назву радіостимуляції, а інтервал доз, у межах якого спостерігається ефект стимуляції, іменують інтервалом стимулюючих доз, а іноді - «малими дозами», хоча з погляду уявлень про радіаційно-хімічні реакції ці дози не такі вже й малі.
Радіостимуляцію виявляють не тільки під час опромінення насіння, але і за впливу іонізуючих випромінювань на цибулини різних рослин, інші органи вегетативного розмноження, наприклад, живці та сіянці.
Форми прояву радіостимуляції, будучи пов’язаними з прискоренням ростових процесів, можуть бути неоднаковими: прискорення проростання насіння, підвищення польової схожості, збільшення галуження, прискорення укорінення живців, підвищення насінної продуктивності рослин, утворення великої вегетативної маси. Відзначається і поліпшення якості продукції.