Альгологія - Костіков І.Ю. - 2009-2013
Розділ 15. Жовтозелені водорості – Xanthophyta
Відділ нараховує біля 600 видів одноклітинних, багатоклітинних та неклітинних сифональних водоростей, що мешкають у прісних континентальних водоймах, ґрунтах та в аерофітоні. Достовірні викопні рештки відомі з тріасових відкладів мезозою (біля 240 млн. років тому), ймовірні - з кам'яновугільного періоду (300-340 млн. років тому). Тобто виникнення жовтозелених водоростей у часі приблизно співпадає з часом появи голонасінних рослин. Більшість жовтозелених водоростей - організми мікроскопічні, і поширені майже виключно у прісних водоймах та ґрунтах.
Жовтозелені водорості належать до трубчастокристних організмів з філи страменопілів, і подібно до інших фотоавтотрофних страменопілів мають пластиди, представлені вторинно симбіотичними родопластами.
Морфологічно жовтозелені водорості дуже нагадують представників відділу зелених водоростей. За наявністю оболонок, основу яких складають пектинові речовини та за відсутністю бурого пігменту фукоксантину жовтозелені водорості схожі із евстигматофітовими, з якими їх об'єднували до 1970 р. Проте Xanthophyta відрізняються від Eustigmatophyta іншим планом будови фоторецепторного апарату монадних клітин, переважанням ксантофілів діатоксантинового ряду над ксантофілами лютеінового ряду, наявністю у зооспор зв'язку ядра з хлоропластною ендоплазматичною сіткою.
На молекулярному рівні жовтозелені водорості виявились більш спорідненими з бурими, ніж з евстигматофітовими водоростями.
Систематичні ознаки відділу
Пігменти та запасні поживні речовини
Біохімічними ознаками відділу є наявність хлорофілів а та с, β-каротину. Переважаючими каротиноїдами є ксантофіли діатоксантинового ряду - діатоксантин та діадіноксантин. У значно менших кількостях у жовтозелених водоростей виявлені специфічні ксантофіли гетероксантин та вошеріоксантин, а також ксантофіли лютеїнового ряду, зокрема неоксантин. Хлоропласти жовтозелених водоростей забарвлені у жовтувато-зелений колір без коричневих відтінків, оскільки типовий для багатьох водоростей-страменопілів темно- жовтий пігмент фукоксантин відсутній.
Продукт асиміляції - хризоламінарин, що завжди відкладається позапластидно, і тому хлоропласти жовтозелених водоростей при оптичній мікроскопії виглядають гладкими та шовковистими, без ознак гранульованості. При старінні у клітинах може накопичуватися олія.
Цитологічні ознаки
Клітинні покриви у жовтозелених водоростей представлені пектиновою або целюлозно-пектиновою оболонкою, яка у деяких представників просякується солями заліза або марганця. Клітинна оболонка може бути суцільною або складатися з двох половинок, що насуваються одна на одну.
У видів роду Vaucheria оболонка крім пектину містить також мікрофібрили целлюлози. Монадні клітини жовтозелених водоростей вкриті лише плазмалемою.
Ядерний апарат типово евкаріотичний. Особливістю ядра є те, що зовнішня мембрана ядерної оболонки переходить у зовнішню мембрану хлоропластної ендоплазматичної сітки, і, таким чином, ядро та хлоропласт у представників Xanthophyta утворюють єдиний структурний комплекс (рис. 15.1). У просторі між зовнішньою та внутрішньою мембранами можуть утворюватися мікротубулярні частини ретронем - тричленних джгутикових мастигонем.
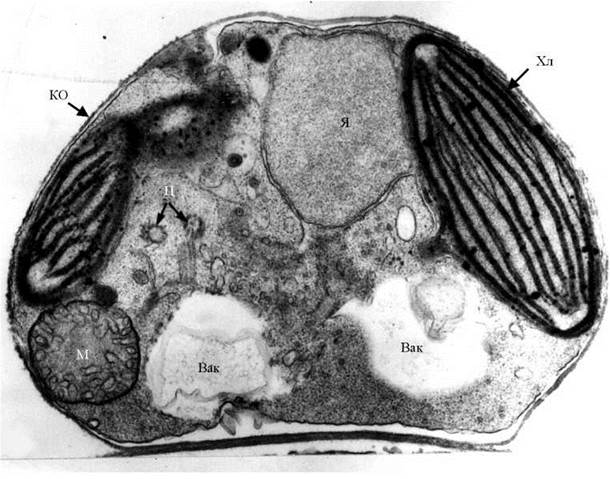
Рис. 15.1. Будова клітини жовтозеленої водорості (на прикладі Heterococcus): КО - клітинна оболонка; Хл - хлоропласт, у якого зовнішня мембрана хлоропластної ендоплазматичної сітки переходить у зовнішню мембрану ядерної оболонки; Я - ядро; М - мітохондрія з трубчастими кристами; Ц - центріолі; Вак - вакуоль (фото А. Массальського).
Мітоз у жовтозелених водоростей закритий, центрами організації веретена є центріолі (рис. 15.2). Зазвичай на полюсах ядра, що ділиться, біля центриолей розташовується комплекс Гольджі.
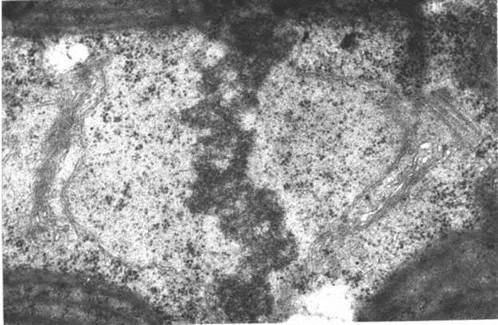
Рис. 15.2. Мітоз у жовтозеленої водорості Xanthonema. Метафаза. Видно інтактну ядерну оболонку, хромосомну платівку, центриоль (фото А. Массальського).
У зооспор після завершення періоду активного руху при першому поділі ядра функції центріолей виконують базальні тіла джгутиків.
Фотосинтетичний апарат. Хлоропласти у жовтозелених водоростей представлені вторинно-симбіотичними пластидами родофітного типу. Кількість хлоропластів у клітині жовтозелених водоростей варіює у широких межах - від одного-десяти у кокоїдних та нитчастих форм до кількох тисяч у сифональних представників. Хлоропласти переважно займають пристінне положення.
Хлоропластна оболонка має чотири мембрани, причому дві зовнішні мембрани утворюють хлоропластну ендоплазматичну сітку, яка переходить у ядерну оболонку. Між зовнішніми та внутрішніми мембранами хлоропласта є перипластидний простір (рис. 15.3).
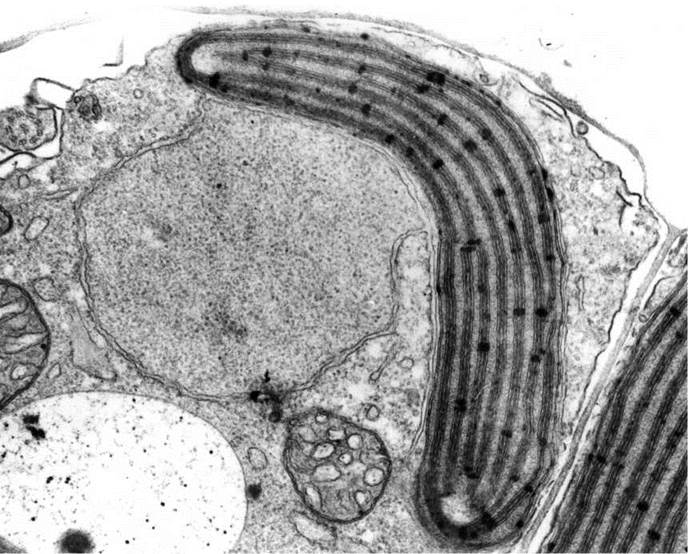
Рис. 15.3. Фрагмент протопласту молодої клітини жовтозеленої водорості Heterococcus. У хлоропласті добре розрізняються тритилакоїдні ламели, оперезуюча ламела, генофор (на полюсах пластиди), а також хлоропластна ендоплазматична сітка з перипластидним простором, ядерна оболонка, від якої відшнуровуються пухирці з зачатковими ретронемами, аксонема джгутика, мітохондрії з трубчастими кристами, вакуоля (фото А. Массальського).
У просторі між мембранами хлоропластної ендоплазматичної сітки, подібно до міжмембранного простору ядра, утворюються тубулярні частини ретронем.
Тилакоїди групуються по три у кілька ламел і зазвичай мають оперезуючу ламелу. Така ламела наявна у більшості представників відділу, проте відомі види, у яких пластида позбавлена оперезуючої ламели (наприклад, з роду Bumilleria). Інколи у пластиді є голий піреноїд, що не пронизаний тилакоїдами.
У зооспор багатьох жовтозелених водоростей у хлоропласті розташовується стигма. Вона утворена одним шаром осмієфільних глобул, притиснутих до поверхні внутрішньої мембрани оболонки хлоропласту. Стигма розміщується поблизу парабазального тіла короткого джгутика.
Хлоропластна ДНК-генофор - компактно зосереджена на полюсах пластиди, і містить 100-200 тис. пар нуклеотидних основ.
Мітохондрії поодинокі або численні, їх профілі на зрізах мають трубчасті кристи. Мітохондрії часто щільно асоційовані з іншими двомембранними органелами - хлоропластами або ядром.
Джгутиковий апарат. Монадні стадії у жовтозелених водоростей представлені зооспорами та гаметами, і на передньому кінці клітини або трохи збоку мають два джгутики нерівної довжини. Довший джгутик пірчастий, і вкритий ретронемами. Утворення ретронем відбувається у пухирцях, що відчленовуються від ядерної мембрани або хлоропластної ендоплазматичної сітки. Короткий джгутик гладенький, і зазвичай виконує функцію керма. При основі короткий джгутик має потовщення - парабазальне тіло, яке є фоторецептором. Стигма у Xanthophyta розташовується в хлоропласті, і виконує функцію ширми, яка з одного боку затіняє фоторецептор. Короткий джгутик може бути редукованим, проте його базальне тіло зберігається. Перехідна зона джгутика містить спіралеподібну структуру.
Від базальних тіл джгутиків відходять три мікротубулярних кореня, кожний з яких складається з трьох-чотирьох мікротрубочок, та один поперечносмугастий мікрофібрилярний корінь - ризопласт, що з'єднує базальні тіла з ядром.
Відхилення від типового плану будови мають зооспори сифональної водорості Vaucheria. Вони великі, багатоядерні; вся поверхня зооспори рівномірно вкрита парами майже однакових за довжиною гладеньких джгутиків. Вважають, що такі сифональні багатоджгутикові зооспори є наслідком злиття у спорангії багатьох молодих дводжгутикових зооспор. Таку об'єднану зооспору називають синзооспорою.
Вакуолярний апарат представлений вакуолями з клітинним соком, що відмежовуються від цитоплазми мембраною - тонопластом. У монадних стадій - зооспор та гамет - біля базальних тіл джгутиків, як правило, є дві скоротливі вакуолі.
Типи морфологічних структур тіла
Більшість жовтозелених водоростей - це кокоїдні, нитчасті, гетеротрихальні або сифональні організми. Крім того, в межах відділу було описано кілька видів з монадною та гемімонадною будовою. Проте слід відмітити, що при спеціальних електронно-мікроскопічних та молекулярно- таксономічних дослідженнях кількох таких представників, всі вони виявилися водоростями з інших відділів - переважно з Chrysophyta та Dictyochophyta. Таким чином, існування у межах Xanthophyta водоростей з монадним та гемімонадним типом морфологічної структури тіла сьогодні ставиться під сумнів.
Розмноження та життєві цикли
Жовтозелені водорості, за винятком одного роду, розмножуються лише нестатевим шляхом: поділом клітин надвоє, фрагментацією таломів або колоній, апланоспорами, зооспорами або за допомогою акінет.
Статевий процес представлений лише оогамією і напевне відомий тільки у видів роду Vaucheria. У цих водоростей життєвий цикл диплофазний, з гаметичною редукцією і без зміни поколінь.
Система відділу
Відділ включає один клас - Xanthophyceae. Згідно з молекулярними даними, еволюція жовтозелених водоростей відбувалась у напрямку переходу від сифональних форм з необмеженим ростом до сифональних з обмеженим ростом, далі - від багатоядерних до одноядерних (кокоїдних, нитчастих та гетеротрихальних). Одноядерні жовтозелені в свою чергу диференціювались за будовою клітинної оболонки на водорості з суцільними оболонками та водорості з двостулковими оболонками. Ці напрямки розвитку знайшли своє відображення у системі порядків класу Xanthophyceae, яка включає чотири основних порядки: Vaucheriales, Botrydiales, Mischococcales та Tribonematales (рис. 15.4).
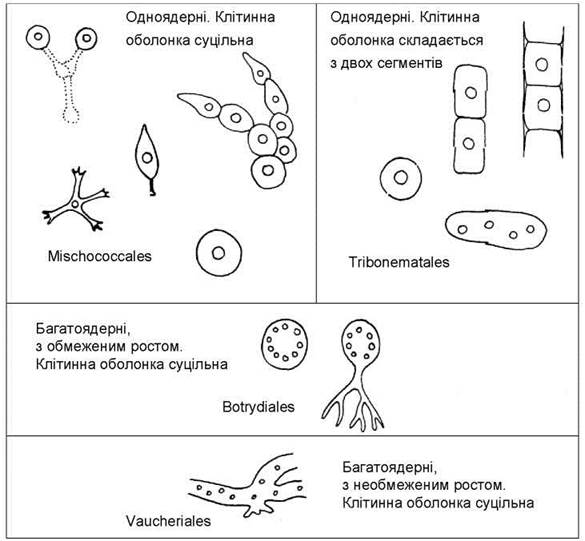
Рис. 15.4. Система Xanthophyta на рівні порядків.
Порядок Vaucheriales об'єднує види з сифональним типом структури, що мають ниткоподібні таломи з необмеженим ростом та суцільні целюлозно- пектинові оболонки (рис. 15.5). Нестатеве розмноження відбувається за допомогою синзооспор. Всі представники порядку можуть розмножуватись статевим шляхом, статевий процес - оогамія. Сперматозоїди мають два джгутики майже рівної довжини, поверхня яких позбавлена ретронем. Порядок містить лише один рід - Vaucheria, до якого входить біля 60 видів. Види роду мешкають на вологому ґрунті, та у солонуватих й прісних водоймах.
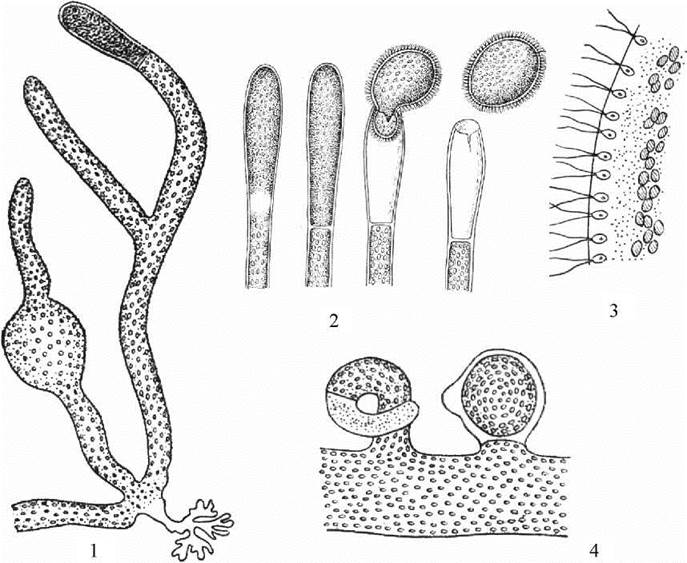
Рис. 15.5. Vaucheria: 1 - зовнішній вигляд талому; 2 - утворення та вихід синзооспори зі споранігію; 3 - фрагмент синзооспори; 4 - фрагмент талому зі статевими органами (ліворуч - антеридій, праворуч - оогоній).
Таломи Vaucheria мають вигляд довгих сланких розгалужених трубок, які позбавлені поперечних перегородок і здатні до необмеженого росту. До ґрунту трубки прикріплюються за допомогою коротких розгалужених ризоїдів. У центральній частині трубок розташовується вакуоль з клітинним соком, між нею та оболонкою - цитоплазма. Безпосередньо під клітинною оболонкою знаходиться велика кількість дрібних дископодібних хлоропластів, між хлоропластами та вакуолею - численні дрібні ядра.
При нестатевому розмноженні деякі верхівкові ділянки талому відокремлюються поперечними перегородками і перетворюються на зооспорангії. В кожному зооспорангії утворюється лише одна гола синзооспора, вкрита парами майже рівних джгутиків. Під базальними тілами кожної пари джгутиків розташовується одне ядро. Після дозрівання синзооспори оболонка спорангію розривається, синзооспора звільняється; після деякого періоду активного руху вона осідає на субстрат і проростає у новий трубчастий сифональний талом.
Статеві органи - антеридії та оогонії - утворюються на одному й тому ж (в однодомних) або на різних (у дводомних) таломах як бічні інтеркалярні вирости, що, як і зооспорангії, відокремлюються від основної частини талому поперечною перегородкою. Після утворення перегородки у гаметангіях відбувається мейоз.
Оогонії вошерії яйцеподібні; в кожному оогонії утворюється по одній яйцеклітині. Антеридії мають вигляд трубчастих гачків. В кожному антеридії утворюється велика кількість сперматозоїдів. Цікаво, що чоловічі гамети, на відміну від синзооспор, мають два латеральні джгутики нерівної довжини, причому довший джгутик несе ретронеми. Загалом будова сперматозоїдів вошерії дуже схожа з будовою сперматозоїдів бурих водоростей.
При статевому процесі сперматозоїди через розрив оболонки антеридія виходять у зовнішнє середовище і підпливають до оогонію. Далі один зі сперматозоїдів через тріщину в оболонці оогонію проникає до яйцеклітини і запліднює її. Зигота одягається товстою оболонкою і переходить в стан спокою. По завершенні періода спокою зигота проростає у новий трубчастий сифональний талом.
Порядок Botrydiales об'єднує сифональні водорості з обмеженим ростом та суцільною пектиновою оболонкою. Таломи ботридіальних можуть мати різноманітну, але не ниткоподібну форму. Зооспори дводжгутикові, гетероконтні, довший джгутик несе ретронеми. Інколи короткий джгутик редукований. Статевий процес у ботридіальних достовірно не виявлений. Характерними представниками порядку є роди Botrydium та Botrydiopsis.
Види Botrydium пристосовані до життя на вологому ґрунті. Їх таломи складаються з системи дихотомічно розгалужених безбарвних ризоїдів, за допомогою яких водорость закріплюється у ґрунті, та розширеної забарвленої надземної частини, діаметр якої досягає 1-2 мм (рис. 15.6: 1). В надземній частині під оболонкою знаходяться численні дрібні дископодібні хлоропласти, під якими розташовується багато ядер. При наявності крапельно-рідкої вологи водорість розмножується дводжгутиковими зооспорами. За посушливих умов протопласт стискається, втягується в ризоїдальну частину і розпадається на одно- або багатоядерні апланоспори. Апланоспори вкриваються міцною, часто - товстою шаруватою оболонкою, і переходять у стан криптобіозу. При настанні сприятливих умов одноядерні апланоспори проростають безпосередньо у нові таломи, а багатоядерні апланоспори перетворюються на зооспорангії з дводжгутиковими зооспорами.
У роду Botrydiopsis вегетеативні клітини сферичні, можуть досягати 0.1 мм у діаметрі (рис. 15.6: 2-4). У периферичному шарі цитоплазми розташовується багато дрібних пристінних дископодібних хлоропластів, під ними - до кількох десятків дрібних ядер. Центр клітини зайнятий вакуолею з клітинним соком. При наявності крапельно-рідкої вологи водорость розмножується за допомогою зооспор; а при її відсутності замість зооспор утворюються одноядерні апланоспори. Види цього мешкають переважно у ґрунтах лісових та тундрових фітоценозів, а також у полярних та високогірних пустелях.
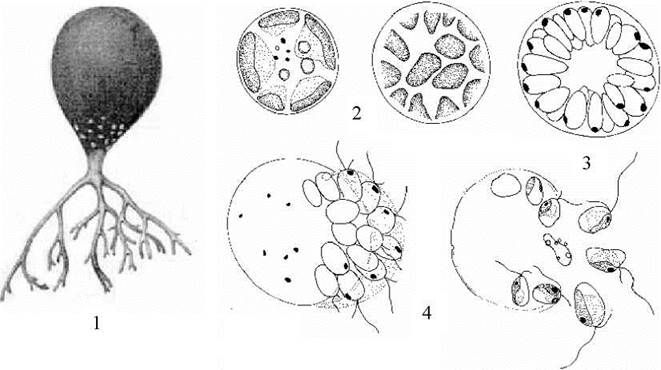
Рис. 15.6. Ботридіальні водорості: 1 - зовнішній вигляд талому Botrydium; 2-4 - Botrydiopsis (2 - вегетативні клітини; 3 - зооспорангій; 4 - вихід зооспор).
До порядку Mischococcales належать одноядерні водорості з суцільними пектиновими оболонками. Примітивніші місхококальні мають кокоїдну будову, складніші - гетеротрихальну.
Найпростішими представниками порядку є види роду Pleurochloris (рис. 15.7: 1-3). Цей рід досить часто трапляється у ґрунтах. Для нього характерні поодинокі сферичні клітини, що нагадують апланоспори ботридіопсіса. Види роду Pseudostaurastrum населяють планктон і мають тетраедричні клітини з довгими відростками, завдяки яким водорість легко утримується у товщі води (рис. 15.7: 4). У перифітонної водорості Characiopsis клітини полярні, і прикріплюються до підводних субстратів за допомогою коротких чи довгих ніжок, що утворені клітинною оболонкою (рис. 15.7: 5).
Представником, що має кокоїдний тип структури і утворює колонії, є рід Mischococcus. У цієї водорості клітини розташовуються на верхівках деревоподібно розгалужених слизових трубок, які прикріплюються до різноманітних підводних субстратів (рис. 15.7: 6-7).
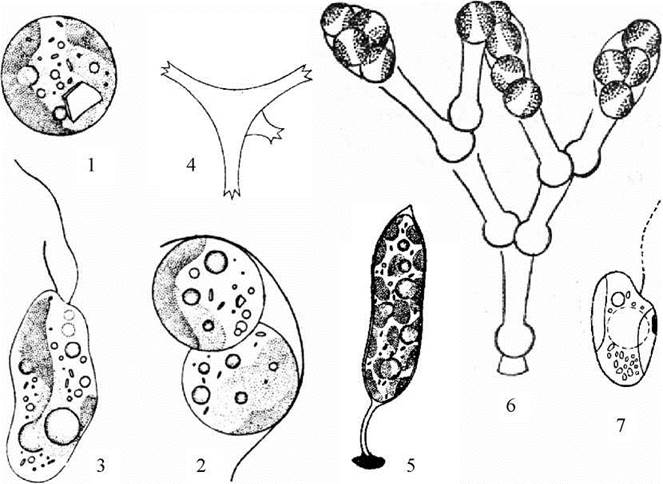
Рис. 15.7. Одноклітинні представники порядку Mischococcales: 1-3 - Pleurochloris (1 - вегетативна клітина; 2 - вихід апланоспор; 3 - зооспора); 4 - Pseudostaurastrum; 5 - Characiopsis; 6-7 - Mischococcus (6 - колонія; 7 - зооспора).
В порядку Mischococcales існують види, що здатні розмножуватись не лише зоо- та апланоспорами, але й шляхом вегетативного поділу клітин (рис. 15.8). Поява у частини місхококальних водоростей вегетативного поділу обумовила виникнення в межах порядку водоростей з простими пластинчастими таломами (наприклад, Chloropedia), і численної групи видів з гетеротрихальним типом структури (Heterococcus). Багатоклітинні місхококальні водорості мешкають майже виключно в наземних умовах: переважно у ґрунтах, рідше - на корі дерев та вологих скелях.
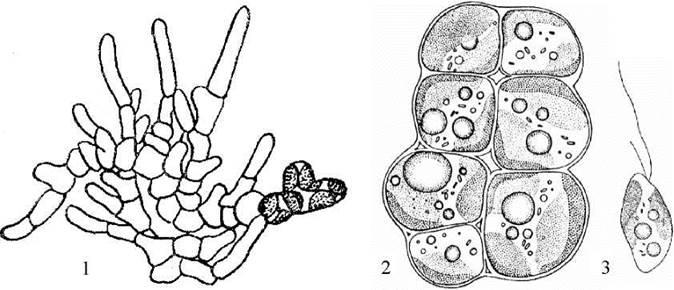
Рис. 15.8. Багатоклітинні представники Mischococcales: 1 - Heterococcus; 2, 3 - Chloropedia (1, 2 - таломи, 3 - зооспора).
Порядок Tribonematales об'єднує водорості, що мають двостулкові клітинні оболонки. У межах порядку представлені одноклітинні та нитчасті організми, причому в обох цих групах відомі представники як з одноядерними, так і з багатоядерними клітинами. За молекулярними даними, порядок відокремився від ботридіальних водоростей, і є сестринським по відношенню до місхококальних.
Серед одноклітинних трибонематальних найпоширенішим є рід Bumilleriopsis, види якого мешкають у ґрунтах та невеличких, переважно ефемерних прісних водоймах. Клітини у Bumilleriopsisвидовжені, ковбасоподібні, вкриті оболонкою, що складається з двох рівних або нерівних сегментів (рис. 15.9: 1-3). Двостулковість оболонки добре простежується при розмноженні водорості - при звільненні з материнської клітини автоспор або зооспор, або після обробки клітин їдким калієм (в останньому випадку оболонка дещо набрякає і стає помітною зона з'єднання двостулкових сегментів). В пристінному шарі цитоплазми вегетативних клітин розташовується кілька дископодібних хлоропластів, в центрі - від одного до чотирьох-восьми ядер, краплини олії та зернятка хризоламінарину. Водорость розмножується зоо- та автоспорами. Інколи при затримці виходу автоспор з материнської клітини Bumilleriopsis утворює агрегати, що нагадують короткі дво- або чотирьохклітинні нитки.
Прикладом виключно одноядерної у вегетативному стані водорості є рід Diachros (рис. 15.9: 4-5). Клітини цієї водорості дрібні, сферичні, і нагадують автоспори Bumilleriopsis. Розмноження відбувається за допомогою автоспор, у яких інколи спостерігаються дві пульсуючі вакуолі. Види цього роду мешкають у кислих ґрунтах та у невеличких кислих водоймах.
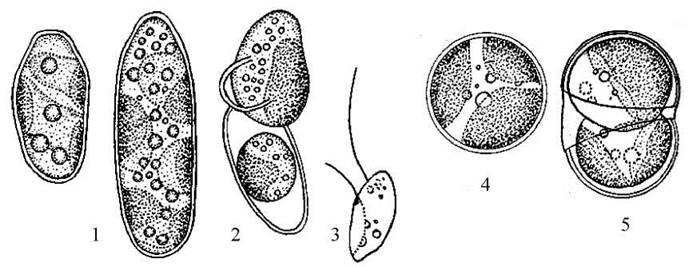
Рис. 15.9. Одноклітинні представники порядку Tribonematales: 1-3 - Bumilleriopsis; 4-5 - Diachros (1, 4 - вегетативні клітини; 2, 5 - вихід апланоспор; 3 - зооспора).
Прикладами поширених нитчастих трибонематальних водоростей є роди Xanthonema та Tribonema.
Водорості з роду Xanthonema мешкають переважно у грунтах. Таломи мають вигляд простих ниток, утворених циліндричними клітинами. Кожна клітина вкрита двостулковою оболонкою (рис. 15.10). У зоні контакту двох сусідніх клітин їх суміжні сегменти щільно або нещільно з'єднуються між собою. В цитолазмі знаходиться від одного до кількох хлоропластів, до яких з внутрішнього боку щільно притиснуті мітохондрії. Центр клітини займають від одного до чотирьох ядер, з'єднаних хлоропластною ендоплазматичною сіткою з пластидою. Біля кожного ядра знаходиться один-два комплекси Гольджі.
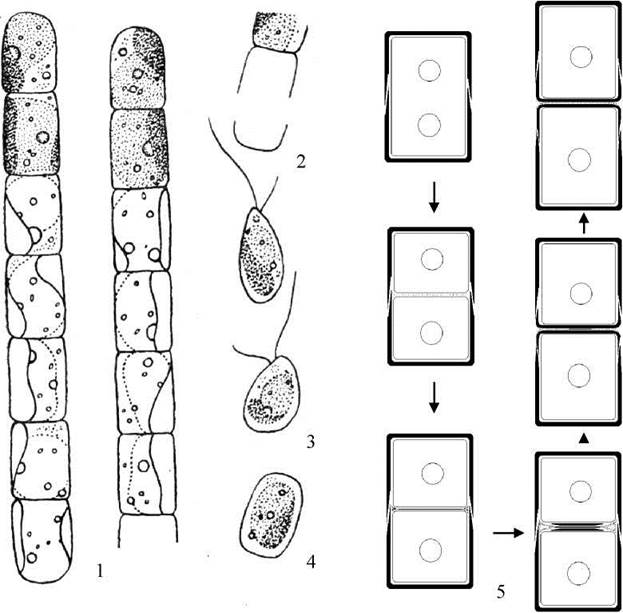
Рис. 15.10. Xanthonema: 1 - зовнішній вигляд ниток; 2 — розкривання двостулкових сегментів при виході зооспор; 3 - зооспори; 4 - апланоспора; 5 - схема послідовних стадій утворення сегментів клітинної оболонки.
Спеціальні електронно-мікроскопічні дослідження дозволили з'ясувати процес утворення клітинних оболонок. Зокрема, було встановлено, що після завершення поділу ядра та протопласту, в простір між плазмалемами двох дочірніх клітин секретується матеріал, що утворює з'єднувальну міжклітинну платівку. Далі у зону платівки до плазмалем дочірніх клітин у пухирцях, похідних від комплексу Гольджі та ендоплазматичної сітки, транспортується матеріал майбутнього сегменту клітинної оболонки. Цей матеріал виводиться у простір між з'єднувальною платівкою та плазмалемою, і полімеризується, утворюючи новий сегмент оболонки дочірньої клітини. Сегмент росте від центру до периферії, одночасно із збільшенням довжини дочірньої клітини. Міжклітинна платівка при цьому зменшується у розмірах, поступово деполімеризуючись у напрямку від периферії до вісі клітини. Якщо процес деполімеризації припиняється, то залишки міжклітинної платівки утримують дочірні клітини разом. Якщо міжклітинна платівка повністю руйнується, то дочірні клітини відокремлюються одна від одної, і відбувається фрагментація талому.
Нестатеве розмноження у Xanthonema здійснюється за допомогою зооспор та апланоспор. Звільнення зооспор та апланоспор здійснюється завдяки розсуванню сегментів оболонки материнської клітини, і тоді нитки закінчуються порожніми напівклітинами, які в оптичному мікроскопі мають вигляд вилочки. При несприятливих умовах вегетативні клітини утворюють додаткові шари клітинної оболонки і перетворюються на акінети.
Види роду Tribonema мешкають переважно у прісних водоймах, інколи утворюючи м'яке світло-зелене жабуріння. Таломи водорості мають вигляд простих нерозгалужених ниток, які на верхівках закінчуються вилочками (рис. 15.11). Процес утворення клітинних оболонок у Tribonema в цілому подібний до Xanthonema, але міжклітинна платівка не утворюється, і матеріал майбутньої клітинної перегородки, що транспортується у мембранних пухирцях, одразу полімеризується з утворенням одного Н-подібного сегменту між плазмалемами двох дочірніх клітин. Як наслідок, суміжні вегетативні клітини міцно з'єднуються між собою, і фрагментація таломів відбувається тільки при розходження двох сегментів однієї клітини. Це обумовлює обов'язкове утворення на верхівках ниток вилочок, і відсутність заокруглених верхівок ниток.
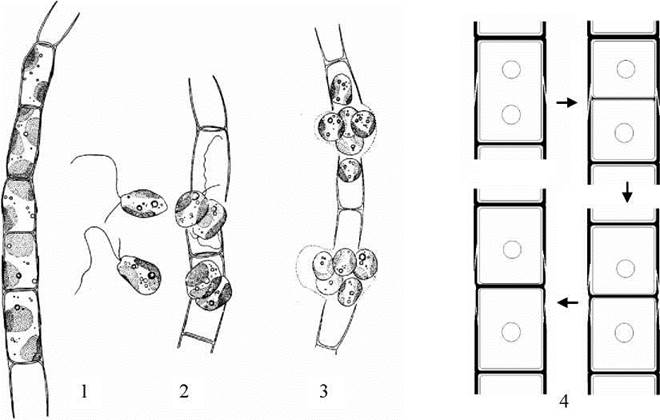
Рис. 15.11. Tribonema: 1 - вегетативна нитка; 2 - вихід зооспор; 3 - вихід апланоспор; 4 - схема послідовних стадій утворення сегментів клітинної оболонки.
На відміну від попереднього роду, клітини у Tribonema у вегетативному стані виключно одноядерні, за винятком стадій утворення репродуктивних клітин - зооспор або апланоспор, та спочиваючих стадій - акінет. Цікаво, що зооспори у цього роду можуть утворюватися різними шляхами: з вегетативних клітин ниток, з апланоспор та з акінет.
Поширення, екологія та значення
Жовтозелені водорості населяють ґрунти та прісні водойми. Найдавніша група жовтозелених, яку представляють види роду Vaucheria, розвиваються переважно на вологому ґрунті - на місці пересохлих калюж, по берегах водойм, і утворюють макроскопічні зелені розростання оксамитової текстури. У схожих умовах ростуть види роду Botrydium. Значна частина жовтозелених водоростей є типовими мешканцями лісових та високогірних ґрунтів. До таких водоростей належать Botrydiopsis, Pleurochloris, Heterococcus, Xanthonema. Ґрунтові Xanthophyta є надзвичайно чутливими до багатьох типів антропогенного забруднення навколишнього середовища, зокрема димо-газового забруднення, забруднень важкими металами, гербіцидами та інсектицидами, детергентами, нафтою та нафтопродуктами, і першим проявом дії таких чинників є зникнення цих водоростей з фітоценозів.
Типовим мешканцем планктону прісних водойм є рід Pseudostaurastrum, а перифітону - Characiopsis та Tribonema. Нитки трибонеми у процесі росту часто відриваються від субстрату, спливають у поверхневі шари води, і там, продовжуючи розмножуватись вегетативним шляхом, утворюють м'які зелені жабуріння. Інші жовтозелені водорості у водоймах зустрічаються зрідка.
Представники відділу Xanthophyta, за винятком родів Vaucheria, Botrydium та Tribonema, у природних умовах майже ніколи масово не розвиваються. Серед Xanthophyta поки що немає об'єктів масової культури і не виявлено шкідливих для людини видів.
Місце Xanthophyta у системі органічного світу
Жовтозелені водорості є типовими тубулокристатами-страменопілами. Цікаво, що саме у жовтозелених водоростей А. Массальським та Г. Лідалем вперше було виявлено тричленні джгутикові волоски, які пізніше отримали назву страменопільних мастигонем (ретронем), від яких, в свою чергу, походить назва "страменопіли".
У 80-х роках ХХ ст. у відділі Xanthophyta на основі електронно-мікроскопічних досліджень Д. Гіббертом та Г. Лідалем було виявлено специфічну групу водоростей, яку на початку 90-х років виділили у самостійний відділ Eustigmatophyta.
У 90-х роках розпочалися роботи по вивченню філогенетичних зв'язків жовтозелених водоростей з іншими відділами на основі молекулярно- генетичних методів, зокрема за результатами секвенування ядерного гену, що кодує 18S субодиницю рибосомальної РНК та хлоропластного гену rbcL, що кодує велику субодиницю рибульозо-1,5-біфосфат-карбоксилази. Ці дослідження підтвердили спорідненість жовтозелених водоростей з іншими водоростями-страменопілами та гіпотезу про походження пластид жовтозелеих водоростей від евкаріотичного пращура з відділу червоних водоростей. За молекулярними даними Xanthophyta виявились ближчими до Phaeophyta, ніж до Chrysophyta та Eustigmatophyta, як це вважалось наприкінці 80-х та початку 90-х років ХХ ст.
Місце в системі
Евстигматофітові водорості належать до підцарства страменопілів царства тубулокристат. За молекулярними ознаками вони близько споріднені із золотистими водоростями, від яких за фенотипічними ознаками відрізняються особливою будовою стигми та піреноїду, наявністю парабазального тіла на довгому джгутику, закритим мітозом, відсутністю оперезуючої ламели, специфічною організацією генофору тощо.