Альгологія - Костіков І.Ю. - 2009-2013
Розділ 14. Евстигматофітові водорості – Eustigmatophyta
У 1970 р. при проведенні електронно-мікроскопічних досліджень Д. Гібберт та Г. Лейдаль звернули увагу на те, що у межах відділу жовтозелених водоростей представлено види з двома відмінними планами ультратонкої будови клітин. Водорості з першим планом будови відповідали "типовим" жовтозеленим водоростям, тоді другий план будови мав риси схожості із золотистими водоростями. Види з другим планом будови було запропоновано виділити у самостійний клас жовтозелених водоростей - Eustigmatophyceae. Пізніше суттєві відміни евстигматофіцієвих водоростей від інших жовтозелених були виявлені також на біохімічному та молекулярному рівнях, внаслідок чого у 90-х роках серед водоростей-страменопілів було встановлено новий відділ - Eustigmatophyta.
Відділ нараховує біля 30 видів одноклітинних, виключно кокоїдних організмів, поширених переважно у ґрунтах та морському планктоні. На думку Т.Нортона зі співавторами (Norton & al., 1996), цей відділ є досить різноманітним і його очікуване видове багатство знаходиться у межах 1000 - 10000 видів.
Систематичні ознаки відділу
Пігменти та запасні поживні речовини
Біохімічними ознаками є наявність хлорофілу а, β-каротину, ксантофілів лютеїнового ряду, серед яких переважає віолаксантин; інші ксантофіли цієї групи - лютеїн, неоксантин, зеаксантин, антераксантин - зустрічаються у слідових кількостях. Зі специфічних ксантофілів у евстигматофітових виявлено вошеріоксантин, який зустрічається також у жовтозелених водоростей з роду Vaucheria. Зелені хлорофіли та жовтий віолаксантин надають хлоропластам евстигматофітових водоростей жовтувато-зеленого забарвлення.
Продукти асиміляції евстигматофітових - це хризоламінарин та олія, а також полісахарид нез’ясованої природи, що відкладається у цитоплазмі навколо піреноїду та у вакуолях.
Цитологічні ознаки
Клітинні покриви представлені пектиновою оболонкою. Оболонка може бути гладкою або орнаментованою і нести потовщення, шипики, бородавки. Монадні клітини евстигматофітових, що представлені зооспорами, вкриті лише плазмалемою (рис. 14.1).
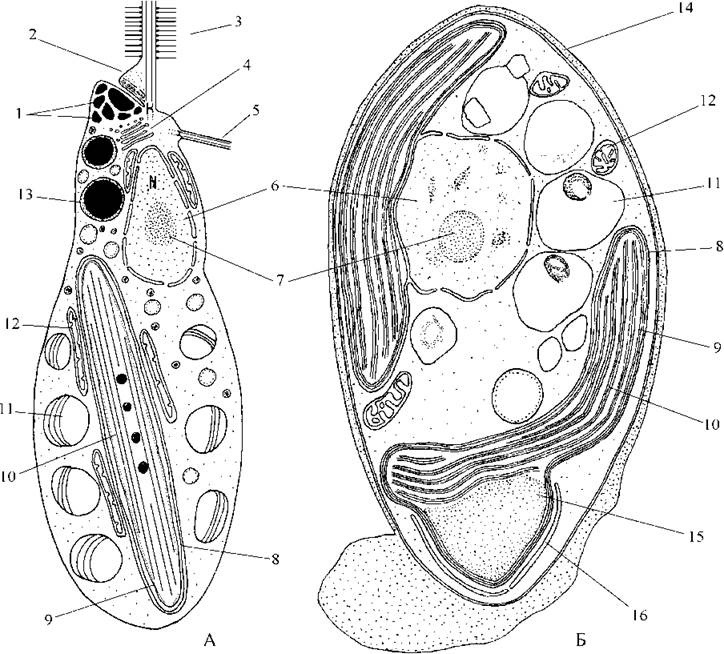
Рис. 14.1. Евстигматофітові водорості. А - зооспора Pseudocharaciopsis, Б - вегетативна клітина Monodopsis. 1 - пігментні глобули стигми; 2 - парабазальне тіло; 3 - довгий джгутик з тричленними мастигонемами; 4 - комплекс Гольджі; 5 - короткий гладенький джгутик; 6 - ядро; 7 - ядерце; 8 - хлоропластна ендоплазматична сітка; 9 - двомембранна оболонка хлоропласта; 10 - тритилакоїдна ламела; 11 - вакуоля з тонкими платівками запасного полісахариду; 12 - мітохондріальний профіль; 13 - краплина олії; 14 - пектинова оболонка; 15 - піреноїд; 16 - платівка запасного полісахариду навколо піреноїду (за Lee, Bold, 1973; Santos, Leedale, 1995).
Ядерний апарат подібний до золотистих водоростей: ядро евкаріотичне, вкрито двомембранною оболонкою. У вегетативних клітин зовнішня мембрана ядра безпосередньо переходить у зовнішню мембрану хлоропластної ендоплазматичної сітки, проте у зооспор зв’язку між ядром та хлоропластною ендоплазматичною сіткою немає.
Мітоз, на відміну від золотистих водоростей, закритий - ядерна оболонка лишається інтактною. Справжні центриолі відсутні, і у монадних клітин їх функцію виконують базальні тіла джгутиків.
Фотосинтетичний апарат. Хлоропласти звичайно поодинокі, переважно розсічені або лопатеві, розташовуються на периферії клітини. Хлоропласт вкритий чотирма мембранами. Два внутрішніх шари утворені мембранами оболонки хлоропласту, два зовнішніх - мембранами хлоропластної ендоплазматичної сітки. Між другою та третьою мембранами є перипластидний простір.
Тилакоїди у стромі хлоропласту зібрані по три, оперезуюча ламела відсутня. У вегетативних клітинах звичайно є піреноїд. Він виступаючий, кутастий, і не пронизаний тилакоїдами. За межами хлоропласту навколо піреноїду відкладаються тонкі платівки асиміляту нез’ясованої природи. Піреноїд завжди відсутній у зооспорах та не виявлений у більшості морських евстигматофітових.
Хлоропластна ДНК, на відміну від Chrysophyta, організована у вигляді числених невеличких агрегатів, що з'єднуються між собою, утворюючи сітчастий розсіяний генофор.
Мітохондріальний апарат поки що досліджений погано. На електронограмах у різних видів спостерігається від двох великих до багатьох дрібних мітохондріальних профілів з трубчастими кристами.
Джгутикові стадії представлені тільки зооспорами. Вони мають два джгутики нерівної довжини - довгий пірчастий локомоторний, що несе два ряди численних мастигонем типової тричленної (страменопільної) будови, та короткий гладенький. Досить часто короткий джгутик редукований і від нього лишається тільки базальне тіло. Тричленні мастигонеми утворюються так само, як і у золотистих водоростей - в інвагінаціях хлоропластної ендоплазматичної сітки та зовнішньої мембрани ядерної оболонки. Перехідна зона джгутиків містить спіралеподібну структуру.
При основі базального тіла пірчастого джгутику знаходиться парабазальне тіло. Його положення зкоординовано з положенням стигми. Остання лежить поза межами хлоропласту і складається з різних за розмірами пігментованих ліпідних глобул, що не оточені плазматичною мембраною. Найбільша глобула огортає з одного боку парабазальне тіло, за нею розташовуються дрібніші глобули. У потовщенні парабазального тіла виявлено електронно-щільну платівкоподібну структуру, яка і є справжнім фоторецептором. Спеціальні дослідження показали, що стигма у евстигматофітових виконує не функцію ширми, як це спостерігається у більшості водоростей, а навпаки, фокусує світлові промені на платівкоподібну структуру парабазального тіла. Саме за унікальною будовою та функцією стигми відділ отримав свою назву - евстигматофітові (рис. 14.2).
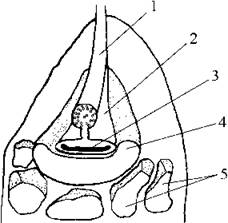
Рис. 14.2. Схема будови фоторецепторного апарату евстигматофітових водоростей: 1 - довгий джгутик, 2 - парабазальне тіло, 3 - платівкоподібна структура, 4 - велика глобула стигми, що огортає парабазальне тіло, 5 - дрібніші глобули зовнішнього шару стигми (за Santos, Melkonian, Kreimer, 1996).
Зооспори евстигматофітових, на відміну від вегетативних кокоїдних клітин, зазавичай позбавлені комплексу Гольджі (виняток - Botryochloropsis similis та Pseudocharaciopsis texensis) та піреноїду, а зовнішня мембрана ядерної оболонки не продовжується у мембрану хлоропластного ендоплазматичного ретикулюму. Коренева система джгутиків евстигматофіцієвих водоростей досліджена погано.
Типи морфологічних структур тіла
Всі відомі евстигматофітові водорості мають тільки кокоїдний тип структури. Більшість видів має поодинокі клітини, деякі представники утворюють слизові (Chlorobotrys) або позбавлені слизу (Botryochloropsis) колонії.
Розмноження та життєві цикли
Розмножуються евстигматофітові тільки за допомогою зооспор та апланоспор. Статеве розмноження не виявлено. Життєві цикли представлені цикломорфозами. Спочиваючі стадії типу цист або акінет у Eustigmatophyta не відомі.
Система відділу
Відділ включає один клас -Eustigmatophyceae, один порядок - Eustigmatalesта чотири родини. Поділ на родини проводиться за морфологічними ознаками - наявністю зооспор, кількістю джгутиків, здатністю утворювати слизові колонії.
Проте результати досліджень гену, що кодує 18S субодиницю рибосомальної РНК свідчать, що у межах відділу існують дві філогенетично відокремлені групи видів. Перша об’єднує ґрунтові та прісноводні евстигматофіцієві водорості, друга - морських пікопланктонних представників. Таким чином, система відділу на рівні родин має штучний характер.
Характерними представниками відділу, що мешкають у ґрунтах є Eustigmatos, Vischeria, Pseudocharaciopsis, Monodopsis, Chlorobotrys, а у морях - Nannochloropsis. Майже всі ці роди раніш входили до складу окремих родів жовтозелених водоростей (наприклад, Eustigmatos - до роду Pleurochloris, Monodopsis - до Monodus, Pseudocharaciopsis - до Characiopsis).
Рід Eustigmatos широко представлений у різних типах ґрунтів всіх регіонів, за винятком тропічних та субтропічних. Найпоширеніший вид - Eustigmatos magnus - легко можна виділити у культуру з лісових, лучних або степових ґрунтів. Вегетативні клітини сферичні, поодинокі, мають великий дво- або трилопатевий хлоропласт, звичайно з добре помітним кутастим піреноїдом, яке раніш називали білковим кристалом. Розмноження у ґрунті та в культурах на агаризованому середовищі відбувається апланоспорами, що утворюються у материнській клітині по 2-4. У водних культурах водорість здатна утворювати одноджгутикові зооспори з досить великою, добре помітною червоною стигмою, що знаходиться біля основи джгутику і не пов’язана із хлоропластом (рис. 14.3).
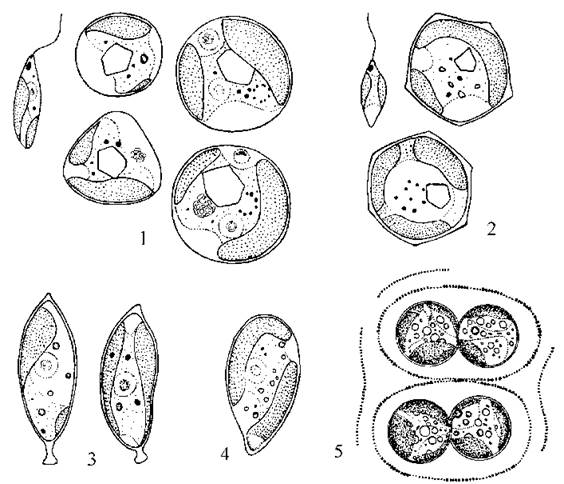
Рис. 14.3. Еустигматофітові водорості: 1 - зооспора та вегетативні клітини Eustigmatos magnus; 2 - зооспора та вегетативні клітини Vischeria helvetica; 3 - Pseudocharaciopsis minuta; 4 - Monodopsis subterranea; 5 - Chlorobotrys regularis (1-4 - ориг., 5 - за Pascher, 1939).
Дуже схожим з Eustigmatos є Vischeria. Її специфічна особливість - наявність кутастих клітин, причому оболонка верхівок кутів потовщена. Види роду є типовими мешканцями гірських ґрунтів.
Pseudocharaciopsis зустрічається як у ґрунтах (переважно - лісових), так і у перифітоні прісних континентальних водойм. Клітини мають веретеноподібну форму, і на одному кінці утворюють невелику ніжку з підошвою, якою прикріплюються до субстрату - піщинок або нитчастих водоростей. Розмноження відбувається апланоспорами та зооспорами, які мають два джгутики нерівної довжини. Найпоширенішим видом є P. minuta.
У роду Monodopsis клітини поодинокі, краплиноподібні, і розмножуються лише апланоспорами. Зустрічаються переважно у наземних біотопах, і інколи (особливо взимку) можуть викликати зелене та жовто-зелене "цвітіння" відмерлих рослинних решіток.
Види роду Chlorobotrys утворюють слизові колонії і розмножуються виключно апланоспорами. Клітини зібрані по дві - чотири у глеокапсоподібні агрегати, що у масі можуть давати макроскопічні розростання світло-зеленого кольору. Представники мешкають переважно у вологих гірських ґрунтах.
Прикладом морської пікопланктонної евстигматофітової водорості є Nannochloropsis. Його клітини дрібні (2-4 мкм у діаметрі), поодинокі, мають один хлоропласт, що позбавлений піреноїду (рис. 14.4). Розмноження відбувається за допомогою апланоспор.
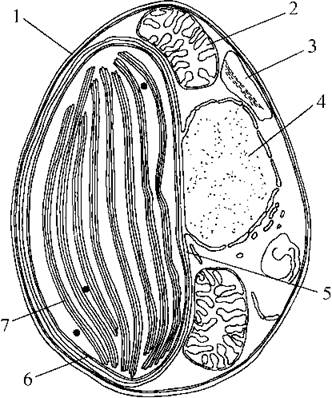
Рис. 14.4. Морська пікопланктонна евстигматофітова водорость з роду Nannochloropsis: 1 - клітинна оболонка; 2 - мітохондріальний профіль; 3 - вакуоля з запасним полісахаридом; 4 - ядро; 5 - хлоропластна ендоплазматична сітка; 6 - оболонка хлоропласту; 7 - тритилакоїдна ламела (схематизовано за Maruyama & al., 1986).
Поширення та значення
Більшість евстигамтофітових водоростей зустрічається у ґрунтах екологічно чистих регіонів, переважно гірських. Представники відділу дуже чутливі до будь-яких форм забруднення ґрунту важкими металами, нафтою та нафтопродуктами, засолення, і при наявності подібних забруднень разом з жовтозеленими водоростями першими випадають із біогеоценозів.
Морські евстигматофітові входять до складу планктонних угруповань відкритих акваторій Світового океану, і разом з іншими мешканцями морського пікопланктону відіграють помітну роль у первинному продукційному процесі.