Альгологія - Костіков І.Ю. - 2009-2013
Розділ 13. Золотисті водорості – Chrysophyta
Золотисті водорості - Chrysophyta - вважаються відносно молодим та нечисленним відділом жовтої пігментної групи. Найдавніші викопні рештки датуються віком біля 240 млн. років (тріасовий період мезозою). На початку мезозою більшість видів відділу вимерла. Сучасну флору Chrysophyta представляють біля 1200 видів, які виникли у палеогені та неогені. Золотисті водорості мешкають переважно у прісних водоймах, і є в основному монадними організмами.
Систематичні ознаки відділу
Пігменти та запасні поживні речовини
Біохімічними ознаками відділу є наявність хлорофілів а та с, β-каротину, та двох груп ксантофілів: по-перше, характерних для жовтої пігментної групи в цілому (фукоксантин, діатоксантин, діадиноксантин, диноксантин), по-друге, ксантофілів лютеїнового ряду (лютеїну, неоксантину, зеаксантину,
антераксантину, віолаксантину). Зелені хлорофіли та жовті ксантофіли (особливо - фукоксантин) надають хлоропластам Chrysophyta яскраво- золотистого відтінку.
Основним продуктом асиміляції є хризоламінарин, додатковими - олія та волютин.
Цитологічні ознаки
Клітинні покриви у Chrysophyta бувають двох типів: монадні та амебоїдні представники переважно голі, водорості гемімонадної, кокоїдної та нитчастої будови вкриті целюлозною або целюлозно-пектиновою оболонкою. Голі клітини можуть бути одягнені лише плазмалемою, нерідко в цьому випадку клітина здатна утворювати псевдоподії; більш міцним покривом є плазмалема, на поверхні якої відкладаються кремнеземові лусочки (див. нижче). Деякі голі клітини знаходяться всередині будиночку з органічних речовин.
Зрідка під плазмалемою знаходяться еджективні структури - дискоболоцисти. Вони являють собою капсулу, утворену цитоплазматичною мембраною; капсула заповнена фібрилярною речовиною, на верхівці капсули розташована згорнута в кільце стрічка. При подразнені плазмалеми стрічка розгортається і вистрілює назовні.
У багатьох колоніальних водоростей під плазмалемою знаходяться також дрібні сферичні слизові тільця. При подразнені клітини вони "вистрілюють" назовні, утворюючи навколо клітини слизову капсулу.
Ядерний апарат. Ядро евкаріотичне, вкрито двомембранною оболонкою, зовнішня мембрана якої безпосередньо переходить у зовнішню мембрану хлоропластної ендоплазматичної сітки (рис. 13.1).
Мітоз відкритий. Справжні центриолі відсутні. Центрами організації веретена поділу є ризопласти. В інтерфазі ризопласт один, розгалужений біля ядерної оболонки, і перед початком поділу реплікується (рис. 13.2).
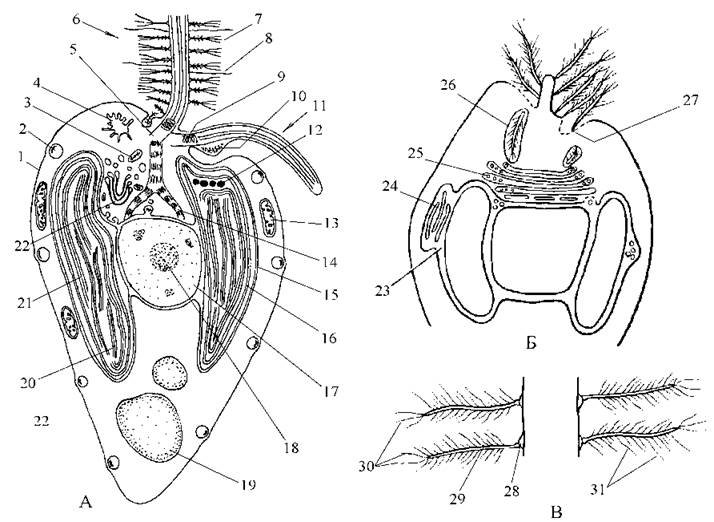
Рис. 13.1. Золотисті водорості. А - схема будови клітини; Б - утворення тричленних мастигонем; В - будова тричленних мастигонем. 1 - плазмалема; 2 - слизове тільце; 3 - пухирець із зачатковою мастигонемою; 4 - скоротлива вакуоля; 5 - базальне тіло довшого джгутика; 6 - довший джгутик; 7 - тричленна мастигонема; 8 - проста мастигонема; 9 - спіралеподібна структура перехідної зони джгутика; 10 - парабазальне тіло; 11 - короткий джгутик; 12 - пігментна глобула стигми; 13 - мітохондріальний профіль із трубчастими кристами; 14 - ризопласт; 15 - мембрани хлоропластної ендоплазматичної сітки; 16 - внутрішні мембрани оболонки хлоропласту; 17 - ядро; 18 - ядерце; 19 - вакуоля з хризоламінарином; 20 - тритилакоїдна ламела; 21 - оперезуюча ламела; 22 - комплекс Гольджі; 23 - простір між мембранами хлоропластної ендоплазматичної сітки, у якому починається синтез тричленних мастигонем; 24 - цистерни хлоропластної ендоплзматичної сітки з матеріалом майбутніх мастигонем; 25 - пухирці комплексу Гольджі, де утворюється зачаткова мастигонема; 26 - везикул із мастигонемою, що транспортується до плазмалеми; 27 - виведення мастигонеми на поверхню плазмалеми; 28 - основа тричленної мастигонеми; 29 - проміжна мікротубулярна частина мастигонеми; 30 - термінальні мікрофібрилярні волоски; 31 - додаткові мікрофібрилярні волоски на поверхні проміжної частини (за Hibberd, 1970; Bouck, 1971).
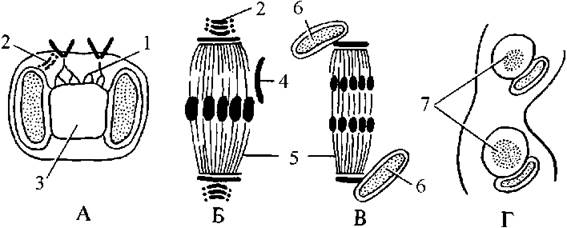
Рис. 13.2. Схема мітозу у золотистих водоростей. А - профаза, Б - метафаза, В - анафаза, Г - телофаза. 1 - ризопласт, 2 - комплекс Гольджі, 3 - ядро, 4 - ендоплазматична сітка, 5 - мікротрубочки веретена поділу, 6 - хлоропласт, 7 - дочірні ядра (за Горбуновою, 1991).
Фотосинтетичний апарат. Хлоропласти у Chrysophyta звичайно поодинокі, розташовуються на периферії клітини. Хлоропластна оболонка чотирьохшарова. Два внутрішніх шара утворені мембранами власне хлоропласту, два зовнішніх є мембранами хлоропластної ендоплазматичної сітки. Між другою та третьою мембранами є перипластидний простір. Зовнішня мембрана хлоропластної ендоплазматичної сітки несе на поверхні 80S рибосоми і відіграє суттєву роль принаймні у процесах формування джгутикових тричленних мастигонем та поверхневих кремнеземових лусочок.
Тилакоїди у стромі хлоропласту зібрані по три, на периферії хлоропласту розташовується оперезуюча тритилакоїдна ламела. Хлоропластна ДНК замкнена у кільце, займає певну ділянку у хлоропласті, тобто генофор концентрований. До його складу входить біля 120 тис. пар нуклеотидів. Інколи у стормі хлоропласту містяться також голі піреноїди та стигма. Стигма у Chrysophytaскладається з одного ряду пігментних глобул, її положення завжди скоординовано з положенням базального тіла короткого джгутика.
Деякі золотисті водорості позбавлені хлоропластів і живляться гетеротрофно. Гетеротрофи поглинають або розчинені у воді органічні сполуки (осмотрофне живлення), або захоплюють часточки їжі псевдоподіями (голозойне живлення).
Мітохондріальний апарат представлений розгалуженим мітохондріоном. На зрізіах мітохондріальні профілі мають трубчасті кристи. Мітохондріальна ДНК має лінійну, а не кільцеподібну організацію і складається приблизно з 40 тис. пар нуклеотидів.
Джгутикові стадії представлені вегетативними клітинами, зооспорами та гаметами. Монадні клітини мають два джгутики нерівної довжини. Довгий джгутик локомоторний, вкритий численними мастигонемами типової тричленної будови, короткий - гладкий або вкритий простими мастигонемами. У багатьох Chrysophyta короткий джгутик редукований і від нього лишається тільки базальне тіло.
На поверхні довгого джгутика пірчасто розташовуються мастигонеми двох типів - тричленні та прості. Тричленні (страменопільні) мастигонеми складаються з базальної частини, прикріпленої до мембрани джгутика, проміжної частини, утвореної мікротрубочками та одного-трьох верхівкових волосков. Крім того, на поверхні проміжної частини знаходиться кілька тонких додаткових ниток (рис. 13.1).
Синтез тричленних мастигонем починається у проміжку між зовнішньою та внутрішньою мемранами ядерної оболонки. У пухирцях, утворених зовнішньою мембраною ядра, зачаткова мастигонема потрапляє до цистерн комплексу Гольджі, де завершується її формування. Далі у пухирцях комплексу Гольджі мастигонема транспортується до плазмалеми при основі джгутика,мембрана пухирця зливається з плазмалемою, залишаючи на поверхні останньої мастигонему, яка поступово переміщується на поверхню джгутику.
Мастигонеми другого типу - т.з. прості - вкривають як короткий, так і довгий джгутики. Вони мають фібрилярну будову і синтезуються безпосередньо на поверхні джгутикової мембрани.
Перехідна зона кожного джгутику містить спіралеподібну структуру. При основі базального тіла короткого джгутику знаходиться потовщення - парабазальне тіло, яке виконує функцію фоторецептора і положення якого скоординовано з положенням стигми.
Джгутикові корені золотистих водоростей складаються з одного поперечно-смугастого кореня - ризопласту та чотирьох мікротубулярних коренів. Ризопласт зв'язує базальні тіла з ядром і побудований зі скоротливого білку центрину.
З базальним тілом кожного джгутику з'єднується по два мікротубулярні корені. Перший корінь складається з трьох мікротубочок, і робить петлю під плазмалемою на передньому кінці клітини. Від цього кореня відходить багато поодиноких мікротрубочок, що направлені до нижнього кінця клітини і створюють каркас цитоскелету. Другий корінь складається з двох мікротрубочок, і також розташовується безпосередньо під плазмалемою. Третій та четвертій корені відходять від базального тіла коротшого джгутику, і поперезують його. Кількість мікротрубочок у цих двох коренів у різних представників різна (рис. 13.3).
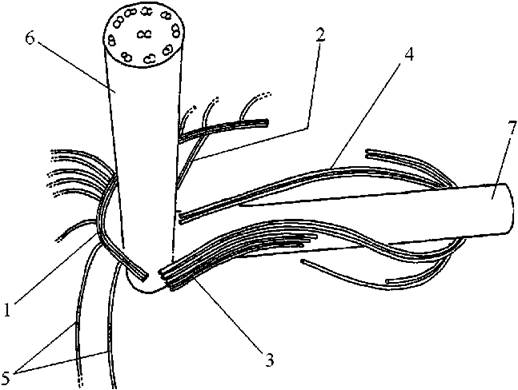
Рис. 13.3. Система мікротубулярних джгутикових коренів золотистих водоростей: 1, 2 - перший та другий корені, що починаються від базального тіла довгого джгутику; 3, 4 - третій та четвертий корені, починаються від базального тіла короткого джгутику; 5 - мікротрубочки, що утворюють цитоскелетний каркас; 6 - довгий джгутик; 7 - короткий джгутик (за Andersen, 1990).
Біля основи джгутиків також розташовується одна-дві скоротливі вакуолі.
Інші органели. Основний продукт асиміляції - хризоламінарин - накопичується у спеціальних одномембранних вакуолях, що розташовуються на задньому кінці клітини.
Типи морфологічних структур тіла
Серед Chrysophyta переважають одноклітинні водорості з монадною та амебоїдною будовою, рідше зустрічаються гемімонадні, кокоїдні та нитчасті представники. Одноклітинні водорості нерідко утворюють колонії та ценобії, багатоклітинні мають вигляд розгалужених або нерозгалужених ниток чи паренхіматозних платівок.
Цисти та їх утворення
Характерною особливістю хризофітових є здатність клітин до утворення ендогенних кремнеземових цист - статоспор. Цисти мають сферичну або еліпсоїдну форму, їх поверхня може бути інкрустована щетинками, шипиками, ребрами. На верхівці цисти знаходиться великий отвір, закритий полісахаридною пробкою; вона звичайно не має кремнезему або містить його лише у слідових кількостях.
Розвиток цисти починається з утворення у цитоплазмі великої цистерни, що отримала назву силіколеми. Вона виникає внаслідок злиття пухирців комплексу Гольджі, і кінець кінцем набуває вигляду двомембранної сфери з отвором на верхівці. Силіколема поділяє цитоплазму клітини на два шари - зовнішній (екстрацистерновий) та внутрішній (інтрацистерновий). У зовнішньому шарі розташовуються скоротливі вакуолі, мітохондрія, частина каналів ендоплазматичної сітки, у внутрішньому - ядро, хлоропласт, комплекс Гольджі, більша частина ендоплазматичної сітки. Між мембранами силіколеми поступово накопичується кремнезем, утворюючи стінки цисти. Далі можуть утворюватися шипи, шипики та щетинки.
По завершенні формування кремнеземової стінки екстрацистернова цитоплазма втягується у цисту, пухирці комплексу Гольджі утворюють полісахаридну пробку, яка з середини закриває отвір цисти. Після цього залишки екстрацистернової цитоплазми, плазмалема материнської клітини та зовнішня мембрана цисти руйнуються, а плазмалемою цисти стає внутрішня мембрана силіколеми. Формування цисти завершується і вона переходить у стан спокою (рис. 13.4).
При проростанні цисти пробка руйнується, протопласт через отвір виходить назовні, і розвивається у нову вегетативну клітину.
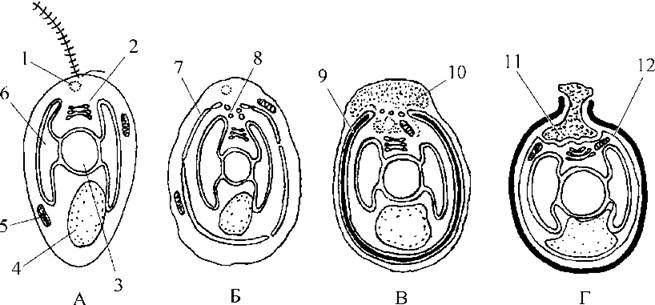
Рис. 13.4. Схема послідовних стадій утворення цисти. А - вегетативна клітина; Б - утворення цистерни силіколеми; В - накопичення кремнезему у силіколемі та початок вгягування у цисту екстрацистернової цитоплазми; Г - зріла циста. 1 - скоротлива ваколя, 2 - комплекс Гольджі, 3 - ядро, 4 - вакуоля з хризоламінаріном, 5 - мітохондріальний профіль, 6 - хлоропласт, 7 - цистерна силіколеми, 8 - пухирці комплексу Гольджі, внаслідок злиття яких утворюється цистерна силіколеми, 9 - кремнеземова стінка цисти всередині силіколеми, 10 - екстрацистернова цитоплазма, що втягується та частково диференціюється у пробку, 11 - пробка, 12 - внутрішня мембрана силіколеми, що стає плазмалемою цисти (за різними авторами).
Розмноження та життєві цикли
Розмножуються Chrysophyta нестатевим та зрідка - статевим шляхами. Нестатеве розмноження у монадних та амебоїдних представників відбувається поділом клітини надвоє у рухливому стані та зооспорами. Крім того, багатоядерні амебоїдні форми (т.з. плазмодіальні водорості) можуть розпадатися на багато одноядерних амебоїдів, кожний з яких в процесі росту та за рахунок мітозів утворює новий плазмодій. Багатоклітинні представники здатні до фрагментації та нестатевого розмноження за допомогою зооспор. Деякі гемімонадні, кокоїдні та нитчасті Chrysophytaрозмножуються також за допомогою апланоспор.
Статевий процес зустрічається зрідка і представлений перважно гологамією, причому клітини під час копуляції з'єднуються задніми кінцями. Спостерігалися також автогамний, ізо- та гетерогамний статевий процеси. Еугамні золотисті водорості мають гаплофазний життєвий цикл з зиготичною редукцією та з особливою стадією двоядерних зиготичних цист (рис. 13.5).
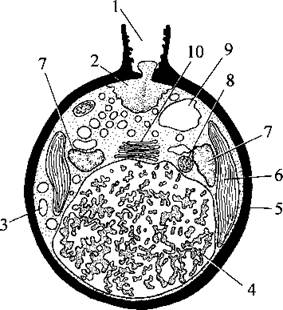
Рис. 13.5. Зиготична циста золотистої водорості Dinobryon: 1 - пора, 2 - пробка, 3 - краплина олії, 4 - вакуоля з хризоламінарином, 5 - стінка цисти, 6 - хлоропласт, 7 - ядро, 8 - мітохондріальний профіль, 9 - скоротлива вакуоля, 10 — комплекс Гольджі (за Горбунова, 1991).
Наприклад, у Dinobryon, Mallomonas, Ochromonas після злиття клітин (плазмогамії) ядра одразу не зливаються. Тому утворюється двоядерна клітина, яка набуває сферичної форми, і перетворюється на цисту характерної для Chrysophyta будови. На відміну від звичайних, ця циста має два різноякісні ядра - жіноче та чоловіче, тобто є дикаріонтичною. Зиготична циста переходить у стан спокою. Після завершення періоду спокою ядра цисти зливаються - відбувається каріогамія і утворюється справжня зигота. Далі диплоїдне ядро зиготи редукційно ділиться, пора відчиняється і через неї виходять одна - чотири гаплоїдні зооспори, які дають початок новим вегетативним клітинам.
Система відділу
Відділ Chrysophyta поділяють на два класи - Chrysophyceae та Synurophyceae. У Chrysophyceae, на відміну від Synurophyceae, клітини не вкриті кремнеземовими лусочками. Ця фенотипічна ознака повністю узгоджується з молекулярно-філогенетичним деревом, що отримано на основі аналізу нуклеотидних послідовностей ядерного гену 18S субодиниці рибосомальної РНК.
До недавнього часу до Chrysophyta включали також клас Prymnesiophyceae. Проте як цитологічні, так і молекулярні дослідження показали, що примнезіофіцієві взагалі не належать до філи страменопілів, і мають бути виділені у самостійний відділ в межах іншого царства - Платикристат.
Клас Chrysophyceae
Об’єднує одноклітинні та багатоклітинні водорості з усіма характерними для відділу типами морфологічних структур - монадним (включаючи амебоїдний та плазмодіальний), гемімонадним, кокоїдним та нитчастим. Ознаками класу є відсутність здатності утворювати кремнеземові лусочки на поверхні плазмалеми. Клас об’єднує прісноводні водорості. У відповідності з морфологічною будовою хризофіцієві поділяються на чотири порядки (табл. 13.1).
Таблиця 13.1. Основні ознаки порядків хризофіцієвих водоростей
Порядок |
Тип морфологічної структури тіла |
Ochromonadales |
монадний |
Chrysamoebales |
монадний, представлений амебоїдним варіантом |
Chrysocapsales |
гемімонадний та кокоїдний |
Phaeothamniales |
нитчастий та гетеротрихальний |
Ochromonadales є найбільш широко розповсюдженими та багатими у видовому відношенні. У порядок входять одноклітинні, колоніальні та ценобіальні види, які мають типову монадну будову. Поширені вони переважно в планктоні чистих прісних водойм. Порядок поділяють на родини та роди за кількістю джгутиків, планами будови тіла (одноклітинний або ценобіальний), покривами (зокрема, наявністю будиночків). Провідними родами порядку є Ochromonas, Chromulina, Dinobryon (рис. 13.6).

Рис. 13.6. Охромонадальні (1-5) та хризамебальні (6-8) водорості. 1 - Ochromonas; 2, 3 - Chromulina: вегетативна клітина (2) та циста (3); 4, 5 - Dinobryon: загальний вигляд колонії (4) та вегетативні клітини після поділу (5) (за Матвієнко, 1965; Starmach, 1968).
Рід Ochromonas характеризується наявністю голих, злегка метаболічних поодиноких монадних клітин з двома нервними джгутиками та одним жовтим хлоропластом. Ochromonas найчастіше зустрічається взимку, у планктоні водойм серед сфагнових болот.
Охромонас вважають найбільш примітивною золотистою водоростю. Цікаво, що деякі види роду здатні утворювати короткі псевдоподії, і таким чином, живитися не тільки фотоавтотрофно, але й голозойно.
Chromulina нагадує попередній рід, проте короткий джгутик редукований, і від нього лишається тільки базальне тіло. Види роду зустрічаються переважно у холодні сезони року у сфагнових болотах.
Клітини видів роду Dinobryon за будовою також подібні до Ochromonas, проте знаходяться всередині будиночків. Останні мають вигляд лійки із сліпо замкненою звуженою основою та розширеною відкритою верхівкою. Кожна клітина прикріплюється до основи будиночку скоротливою базальною ніжкою.
Органічні речовини будиночку - мікрофібрили целюлозні з домішкою амінокислот - синтезуються у комплексі Гольджі, далі транспортуються на поверхню плазмалеми, і там полімеризуються.
При розмноженні протопласт материнської клітини ділиться, утворюючи дві голі дочірні клітини. Вони виходять з будиночку, осідають на його край біля отвору, і далі будують власні будиночки. Внаслідок цього звичайно утворюються деревоподібні дихотомічно розгалужені колонії.
Види роду є типовими мешканцями планктону прісних водойм (найчастіше - озер), і зустрічаються о будь-якій порі року.
Chrysamoebales об’єднує водорості з амебоїдною будовою. Клітини хризамебальних здатні утворювати тонкі нерозгалужені або розгалужені ризоподії, рідше - короткі аксоподії.
Характерний представник - Chrysamoeba (рис. 13.6). Її клітини поодинокі, мають довгі численні ризоподії. У цитоплазмі добре помітно золотистий хлоропласт, вакуолю з хризоламінарином, скоротливі вакуолі. Амебоїдні клітини здатні переходити у короткочасний рухливий стан. При цьому ризоподії втягуються та утворюється один довгий джгутик. За несприятливих умов водорость переходить у пальмелевидний стан, одягаючись слизовою капсулою. Види роду мешкають переважно у планктоні стоячих водойм.
Chrysocapsales - невеликий порядок, до якого належать водорості з гемімонадним, рідше - кокоїдним типами структур. Як правило, клітини хризокапсальних водоростей занурені у колоніальний слиз. Майже всі хризокапсальні є рідкісними водоростями.
Частіше від інших зустрічаються рід Hydrurus - прісноводна бентосна водорість, та рід Chrysostephanosphaera, що мешкає у планктоні невеликих прісних водойм (рис. 13.7). Цікаво, що клітини хризостефаносфери здатні переходити у монадний або амебоїдний стан, і тоді нагадують хромуліну або хризамебу, відповідно. Розмножуються хризокапсальні водорості зооспорами.
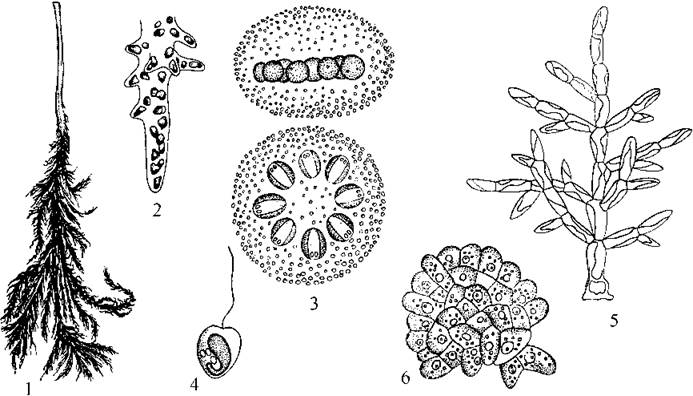
Рис. 13.7. Хризокапсальні (1-4) та феотамніальні (5, 6) водорості. 1, 2 - Hydrurus: загальний вигляд колонії (1) та фрагмент з клітинами (2); 3, 4 - Chrysostephanosphaera: колонія (3) та зооспора (4); 5 - Phaeothamnion; 6 - Phaeodermatium (за Топачевский, Масюк, 1984).
Phaeothamniales - об'єднує невелику кількість багатоклітинних прісноводних видів. Таломи нитчастих феотамніальних одно- або багаторядні, розгалужені або прості. Гетеротрихальні представники мають таломи у вигляді паренхіматозних платівок. Характерними родами є Phaeothamnion (має талом у вигляді однорядної розгалуженої нитки) та Phaeodermatium (талом у вигляді паренхіматозної платівки), що зрідка зустрічаються у чистих гірських річках.
Клас Synurophyceae
Об’єднує одноклітинні, переважно - монадні водорості, плазмалема яких вкрита спіралеподібно розташованими кремнеземовими лусочками. Звичайно ці лусочки добре помітні в оптичний мікроскоп. Кожна лусочка складається з платівки та з'єднаної з нею щетинки. Платівка перфорована дрібними порами, має потовщений край та інколи - дрібні шипика та бородавки. Щетинка приєднана до нижнього краю платівки, і має вигляд порожнистої трубки.
В утворенні платівок та щетинок приймають участь хлоропластна ендоплазматична сітка та пухирці, що є похідними комплексу Гольджі. Так, спочатку від зовнішньої мембрани хлоропластної ендоплазматичної сітки відбруньковується пухирець їх кремнеземом. У цитоплазмі він зливається із пухирцями, що відбрунькувалися від комплексу Гольджі. Утворюється везикул, у якому завершується формування або платівки, або щетинки. Везикул переміщується до поверхні клітини, його мембрана зливається із плазмалемою, і елемент частини лусочки (або платівка, або щетинка) потрапляє на поверхню плазмалеми. Першими виштовхуються щетинки, за ними - платівки. З’єднання щетинок та платівок у нормальну лусоку відбувається вже поза межами клітини.
Клас включає один порядок - Synurales (= Mallomonadales).
Найбагатшим родом класу є Mallomonas. Клітини водорості поодинокі, вільноплаваючі і мають лише один джгутик. На поверхні плазмалеми розташовуються численні черепитчасто розміщені кремнеземові лусочки з довгими щетинками. Цисти також мають додатковий шар лусочок, звичайно з довгими шипами.
Водорості роду Synura утворюють сферичні колонії з багатьох клітин, з’єднаних задніми кінцями. Кожна клітина має два джгутики. Плазмалема вкрита типовими для Synurophyceaeкремнеземовими лусочками, кожна з яких складається з платівки та короткої щетинки.
У деяких видів (зокрема, S. petersenii) виявлено та детально досліджено статеве розмноження. Тип статевого процесу - хологамія. Вегетативні клітини, що виконують функції жіночих гамет, виділяють аттрактивний гормон. Під його дією клітини, що виконують функції чоловічих гамет, залишають колонію, підпливають до жіночої клітини та копулюють. Як наслідок, утворюється зиготична дикаріонтична циста. Її подальший розвиток відбувається за звичайною для золотистих водоростей схемою.
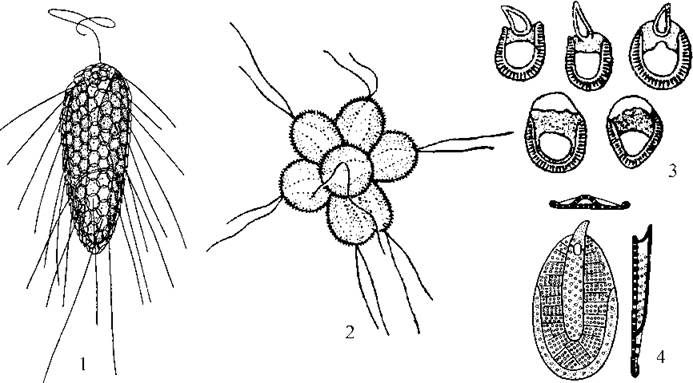
Рис. 13.8. Синурофіцієві водорості. 1 - Mallomonas; 2-4 - Synura: колонія (2), різноманітність лусочок (3); одна лусочка при вигляді у трьох площинах (4) (за Матвієнко, 1965; Горбунова, 1991).
Поширення та значення
Золотисті водорості поширені переважно у прісних водоймах. Як і більшість представників жовтої пігментної групи, хризофітові є холодолюбними організмами, що масово розвиваються або у приполярних широтах, або у холодні пори року у помірних широтах.
Золотисті водорості з кремнеземовими лусочками утворили осадкові силікатні породи - археомонадити. Викопні залишки Chrysophyta разом з залишками інших організмів використовують в геології для визначення віку осадових порід.
Прісноводні Chrysophyta зустрічаються переважно у планктоні чистих водойм (в основному - види Chromulina), значно рідше - у бентосі та перифітоні (Chrysamoebales, Chrysocapsales, Phaeothamniales). У холодні пори року деякі прісноводні представники можуть викликати золотисте або жовто-коричневе "цвітіння" (види родів Chromulina, Mallomonas, Dinobryon, Synura); вода в цьому випадку набуває характерного запаху застарілого риб’ячого жиру. Деякі золотисті водорості використовуються для оцінки якості води.
Місце в системі
Chrysophyta є типовим відділом фотоавтотрофних тубулокристат- страменопілів, і за молекулярними даними, що корелюють з деякими морфологічними та цитологічними ознаками, найбільш споріднений з віділом Eustigmatophyta.