Альгологія - Костіков І.Ю. - 2009-2013
Розділ 12. Рафідофітові водорості – Raphidophyta
До 70 -х років нашого століття рафідофітові водорості під назвою Chloromonadophyceae об'єднували з криптофітовими та динофітовими водоростями у відділі Pyrrophyta або з евгленофітовими у відділі Euglenophyta. Накопичення даних про біохімічні особливості та цитологічну специфіку видів, що входили до Chloromonadophyceae, призвело до виділення цієї групи в самостійний відділ під назвою Chloromonadophyta. Згодом, згідно з вимогами Міжнародного Кодексу Ботанічної номенклатури відділ був переіменований на Raphidophyta. У цьому відділі сьогодні відомо близько 50 видів з 10 родів, усі представники є виключно одноклітинними монадними водоростями, що мешкають у прісних континентальних водоймах та у морях.
Систематичні ознаки відділу
Пігменти та запасні поживні речовини
Біохімічними ознаками відділу є наявність хлорофілів а та с, β-каротину, ксантофілів лютеїнового ряду (віолаксантину, лютеїну). Крім того, у прісноводних рафідофітових присутні діадиноксантин, диноксантин, гетероксантин, вошерієксантин, а у морських форм - фукоксантин. Через це хлоропласти у прісноводних видів забарвлені звичайно у зелений або жовто- зелений колір, а у морських - у жовтий або жовто-коричневий.
Продуктом асиміляції є олія, що накопичується в цитоплазмі1. В окремих випадках дрібні поодинокі краплини олії виявляються також у стромі хлоропласта.
Цитологічні ознаки
Клітини рафідофітових досить великі (30-100 мкм), звичайно дорзовентральної будови, з трикутною глоткою, двома гетероконтними та гетероморфними джгутиками, вузькою поздовжньою борозенкою, що розташовується на вентральному боці клітини.
Клітинні покриви - плазмалема, під якою можуть знаходитись слизові тільця або/та трихоцисти. При подразненні трихоцисти викидають назовні слизові нитки.
Ядерний апарат рафідофітових водоростей має типову евкаріотну будову. Ядро одне, велике, з одним або кількома ядерцями. Над ядерною оболонкою у вигляді широкого ковпачка розташовується т.з. супрануклеарний апарат. Електронно-мікроскопічні дослідження показали, що супрануклеарний апарат складається з кількох комплексів Гольджі, які щільно прилягають до ядерної оболонки. З супрануклеарним апаратом з'єднуються мікрофібрилярні джгутикові корені (рис. 12.1).
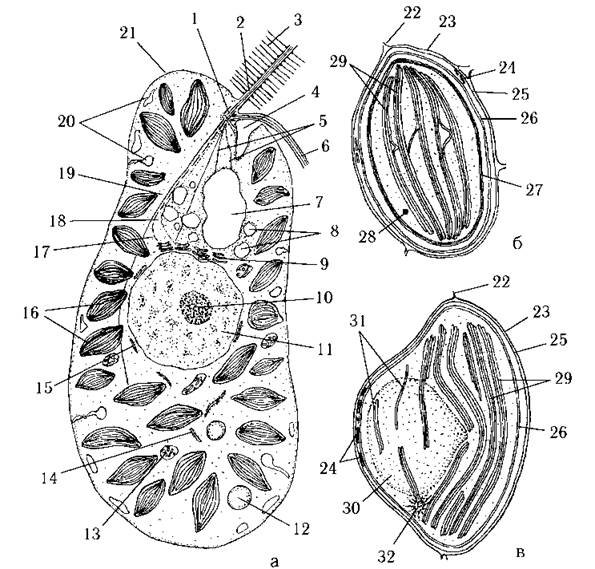
Рис. 12.1. Будова клітини (а) та хлоропласта (б, в) рафідофітових водоростей: 1 - базальні тіла джгутиків, 2 - пірчастий джгутик, 3 - тричленні мастигонеми, 4 - глотка, 5 - мікротубулярні корені, що з'єднують базальні тіла з вакуолярним апаратом, 6 - гладенький джгутик, 7 - велика скоротлива вакуоля, 8 - дрібні скоротливі вакуолі, 9 - один з комплексів Гольджі супрануклеарного апарату, 10 - ядерце, 11 - ядро, 12 - краплина олії, 13 - мітохондрія з трубчастими кристами, 14 - канал ендоплазматичної сітки, 15 - фрагмент навколоядерної цистерни ендоплазматичної сітки, 16 - хлоропласти, 17 - поперечно-смугастий мікрофібрилярний корінь, 18 - гладенький мікрофібрилярний корінь, 19 - мікротубулярний корінь з багатошаровою структурою, 20 - слизові тільця, 21 - плазмалема, 22 - канал ендоплазматичної сітки, що переходить у зовнішню мембрану хлоропласту, 23 - перша зовнішня мембрана хлоропласта, 24 - перипластидний ретикулюм, 25 - друга зовнішня мембрана хлоропласта, 26 - дві внутрішні мембрани хлоропласта, 27 - оперезуюча ламела, 28 - дрібна краплина олії, 29 - тритилакоїдні ламели, 30 - піреноїд, 31 - поодинокі та парні тилакоїди, що пронизують строму піреноїда, 32 - генофор (а, б - прісноводний представник (Vacuolaria), в - морський (Fibrocapsa). Схематизовано за Heywood, 1972, 1977, 1990; Hara, Chihara, 1985).
Мітоз у рафідофітових закритий: ядерна оболонка під час поділу ядра лишається інтактною, а веретено є внутрішньоядерним. Функції центріолей виконують базальні тіла джгутиків.
Фотосинтетичний апарат. Хлоропласти у Raphidophyta дрібні, численні, розташовані на периферії клітини. Кожний хлоропласт вкритий чотиримембранною оболонкою. Дві зовнішні мембраниутворюють хлоропластну ендоплазматичну сітку, яка переходить у канали ендоплазматичного ретикулюму, проте не продовжується у ядерну мембрану. Між двома зовнішніми та двома внутрішніми мембранами хлоропласта є перипластидний простір з перипластидним ретикулюмом.
Тилакоїди групуються по три у кілька ламел. У прісноводних видів хлоропласти позбавлені піреноїдів і мають оперезуючу ламелу. У морських представників піреноїди є, вони напівзанурені і звичайно пронизані двома тилакоїдами, а оперезуюча ламела відсутня. Хлоропластна ДНК-генофор - замкнена у кільце і знаходиться поблизу хлоропластної оболонки.
Мітохондрії розгалужені, утворюють мітохондріон. Мітохондріальні профілі на зрізах мають трубчасті кристи.
Джгутиковий апарат. На передньому кінці клітини або трохи збоку розташовуються два джгутики нерівної довжини. Довший джгутик пірчастий, на кінці звичайно спірально закручений, забезпечує рух клітини. Він вкритий мастигонемами тричленної (т.з. страменопільної) будови. Утворення мастигонем відбувається у цистернах ендоплазматичного ретикулюму. Короткий джгутик гладенький, звичайно виконує функцію керма і лежить у поздовжній черевній борозенці. Парабазальні потовщення на джгутиках та вічко у рафідофітових не виявлені. Перехідна зона джгутика не має спіралеподібної структури.
Від базальних тіл джгутиків відходять дві системи коренів - глоткова та ядерна. Глоткова система складається з трьох-чотирьох мікротубулярних коренів, які розміщуються в цитоплазмі поблизу глотки. Ядерна система коренів включає один мікротубулярний корінь з багатошаровою структурою, та два мікрофібрилярні корені - гладенький та поперечно-смугастий. Мікрофібрилярні корені пов’язують базальні тіла джгутиків зі супрануклеарним апаратом, а мікротубулярний корінь проходить вздовж ядра до заднього кінця клітини.
Вакуолярний апарат представлений однією великою та кількома дрібними скоротливими вакуолями, що виштовхують свій вміст у глотку.
Тип структури тіла
Всі рафідофітові водорості мають монадний тип структури тіла. Клітини злегка метаболічні, переважно дорзовентральні або стиснуті з боків, з поздовжньою борозенкою, від якої у безбарвних форм можуть відходити ризоподії.
Розмноження та життєві цикли
Розмноження відбувається поділом клітини у поздовжньому напрямку у рухливому стані. Перед початком поділу гладенький джгутик звичайно скидається, пірчастий реплікується. Після закінчення поділу кожна клітина має пірчастий джгутик і добудовує гладенький. Зооспори та статеве розмноження у Raphidophyta не відомі.
Життєвий цикл простий, подібний до циклів криптофітових водоростей. При несприятливих умовах відбувається перехід у пальмелевидний стан. Спостерігаються також цисти сферичної форми, які інколи мають отвір, закритий пробкою.
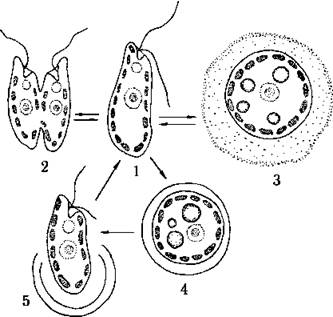
Рис. 12.2. Життєвий цикл рафідофітових водоростей: 1 - монадна вегетативна клітина, 2 - поділ клітини надвоє у монадному стані, 3 - клітина у пальмелевидному стані, 4 - циста, 5 - проростання цисти монадною вегетативною клітиною.
Особливості живлення
Переважна більшість рафідофітових - фотоавтотрофи. Проте деякі представники відділу позбавлені хлоропластів і є облігатними гетеротрофами. Живлення гетеротрофних видів відбувається осмотрофним шляхом. Зустрічаються також види, здатні до голозойного живлення. Вони захоплюють їжу (бактерії, частки детриту, інші водорості) ризоподіями; травлення відбувається у травних вакуолях.
Система відділу
Всі Raphidophyta відносяться до одного класу - Raphidophyceae, та двох порядків - Raphidiales (=Vacuolariales) та Chattonellales. В основу поділу на порядки покладено екологічні особливості, склад додаткових ксантофілів, особливості будови хлоропласта, зокрема - наявність піреноїдів та оперезуючої ламели.
Порядок Raphidiales об’єднує прісноводні фототрофні та гетеротрофні рафідофітові водорості, у яких у складі пігментів переважають діадиноксантин та диноксантин, є оперезуючі тилакоїди і відсутні піреноїди. Поділ на родини та роди відбувається, в першу чергу, за типом живлення, наявністю чи відсутністю трихоцист, розташуванням глотки, формою клітини, здатністю утворювати псевдоподії. Основними родами є Vacuolaria, Goniostomum, Merotrichia, Thaumatomastix, Hyaloselene (рис. 12.3).
Роди Vacuolaria, Goniostomum, Merotrichia об’єднують фотоавтотрофні водорості. У Vacuolaria є слизові тільця, проте відсутні трихоцисти. У Goniostomum та Merotrichia - навпаки, причому у першого джгутики та глотка майже апікальні, а у другого - зміщені на черевний бік. Роди Thaumatomastix та Hyaloselene являють собою гетеротрофні водорості, які живляться як осмотрофно, так і голозойно, захоплюючи частки їжі (детрит, бактерії, інші водорості) за допомогою псевдоподій.
Порядок Chattonellales об’єднує морські автотрофні рафідофітові, у яких головний ксантофіл - це фукоксантин, наявні піреноїди і відсутні оперезуючі тилакоїди. Характерними морськими представниками є роди Chattonella, Heterosigma та Fibrocapsa (рис. 12.4).
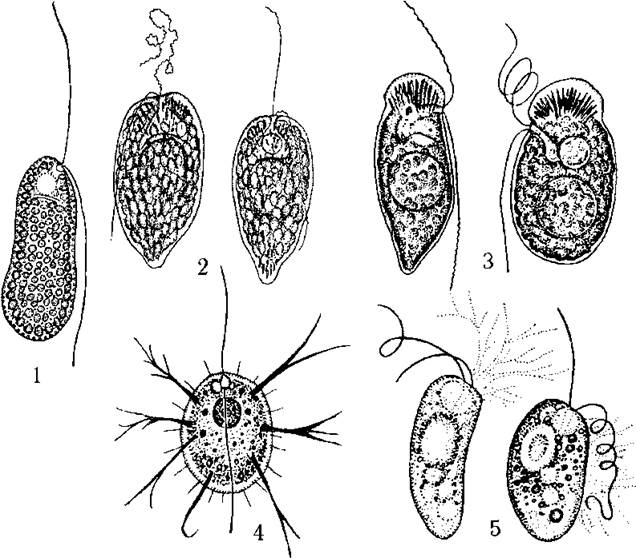
Рис. 12.3. Деякі представники порядку Raphidiales: 1 - Vacuolaria, 2 - Goniostomum, 3 - Merotrichia, 4 - Thaumatomastix, 5 - Hyaloselene (за Матвієнко, Литвиненко, 1977).
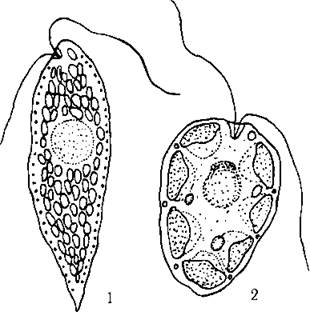
Рис. 12.4. Деякі морські рафідофітові водорості з порядку Chattonellales: 1 - Chattonella, 2 - Fibrocapsa (схематизовано за Hara, Chihara, 1982, 1985; Khan et al., 1996).
Поширення, екологія та значення
Всі рафідофітові водорості - представники планктону прісних водойм та морів. Прісноводні рафідофітові є переважно ацидофілами, і найчастіше зустрічаються у сфагнових болотах навесні та восени. Морські рафідофітові належать до нейтрофілів.
Тривалий час вважалося, що рафідофітові водорості не мають практичного значення. Проте на початку 80-х років з’явилися перші дані про токсичні "червоні припливи" біля узбережжя Японії, спричинені рафідофітовими водоростями Chattonella antiqua та Ch.marina. Ці "цвітіння" спричинили масову загибель риби у рибоводних садках, розташованих у морі, й завдали значної шкоди економіці аквакультури. Пізніше токсичні види були знайдені й серед інших родів морських Raphidophyta.
На теперішній час у рафідофітових виявлено три нейротоксини, гемолітичні токсини та гемаглютинуючі токсини. Навіть при малій кількості клітин (3-10 тис.кл. /мл) рафідофітові порушують координацію руху у риб, спричинюють їх аномальну поведінку, і протягом 50-80 хвилин призводять до загибелі. Найбільшу токсичність рафідофітові мають на стадії активної вегетації - у т.з. фазі логаріфмічного росту.
Місце Raphidophyta у системі органічного світу
Порівняно недавно - ще у 80-і роки - рафідофітові розглядали як таксон з проблематичними філогенетичними зв'язками: деякі автори наближали Raphidophyta до криптофітових водоростей, інші - до евгленофітових. Проте після відкриття страменопільного плану будови джгутикових мастигонем рафідофітові почали розглядати як примітивних страменопілів. Подальші цитологічні дослідження (зокрема, з’ясування плану будови хлоропласту, морфології мітохондріальних крист) підтвердили обгрунтованість такої точки зору.
У 1996 р. у рафідофітової водорості Heterosigma carterae було розшифровано нуклеотидну послідовність ядерного гена, що кодує 18S субодиницю рибосомальної РНК. Це дало можливість залучити рафідофітові до молекулярно-філогенетичного аналізу. На підставі молекулярно-філогенетичних реконструкцій було підтверджено останню точку зору, а також отримано дані, що Raphidophyta найбільш споріднені із золотистими та евстигматофітовими водоростями.
1 Крохмаль, який раніш вважали також характерним для рафідофітових водоростей, був відомий лише у одного роду Monomastix, який зараз вилучений з Raphidophyta і перенесений до зелених водоростей.