Альгологія - Костіков І.Ю. - 2009-2013
Розділ 11. Хлорарахніофітові водорості – Chlorarachniophyta
Відділ Хлорарахніофітових водоростей (Chlorarachniophyta) встановлений у 1984 р. під час дослідження ультратонкої будови Chlorarachnion reptans - амебоїдної водорості, яку раніш включали до відділу Xanthophyta. Цей вид вперше був виділений з культури морського мулу у 20-х роках н.ст. У 1987 р. було описано другого представника відділу - Cryptochlora perforans, виділеного у культуру з відмерлих таломів морських сифональних водоростей. Пізніше було знайдено ще кілька видів з цієї філи. Сьогодні хлорарахніофітові включають чотири роди з шістьма видами.
Біохімічні, а згодом - і молекулярно-біологічні дослідження підтвердили самостійність хлорарахніофітових як таксону вищого рангу.
Систематичні ознаки відділу
Пігменти та запасні поживні речовини
Біохімічними ознаками відділу є наявність хлорофілів а та b, які не маскуються додатковими пігментами. Через це хлоропласти хлорархніофітових забарвлені у зелений колір. Запасною поживною речовиною, подібно до евгленофітових, є парамілон.
Цитологічні ознаки
Клітини хлорарахніофітових амебоїдні й утворюють велику кількість тонких розгалужених ризоподій. Вони з'єднують клітини у сітчасті колонії, які інколи називають плазмодієм. Кількість клітин у колонії коливається від кількох до 150. Кожна клітина колонії здатна до активного руху за допомогою псевдоподій (рис. 11.1).
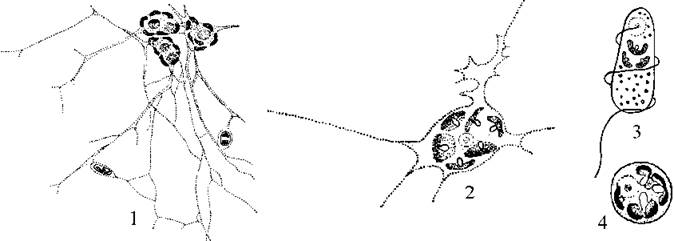
Рис. 11.1. Хлорарахніофітові водорості (на прикладі Chlorarachnion): 1 - колонія; 2 - окрема амебоїдна клітина; 3 - зооспора; 4 - кокоїдна стадія (за Geitler, 1930; Pascher, 1939).
Клітинні покриви представлені лише плазмалемою. Під плазмалемою знаходяться дифузно розміщені трихоцисти.
Ядерний апарат. Клітини хлорарахніофітових одноядерні. Ядро має евкаріотну будову, хромосомну організацію, ядерна ДНК зв’язана з гістонами. Ядро відмежовано від цитоплазми двомембранною оболонкою і містить одне ядерце.
Фотосинтетичний апарат. У клітині міститься кілька пристінних дископодібних дволопатевих хлоропластів, кожний з яких має піреноїд. Хлоропласти вкриті чотирьохмембранною оболонкою. Дві внутрішні мембрани є власними мембранами хлоропласту, дві зовнішні мембрани утворюють хлоропластний ендоплазматичний ретикулюм, звичайно - принаймні з однією великою інвагінацією. Між власними хлоропластними мембранами та мембранами хлоропластного ендоплазматичного ретикулюму є досить великий, т.з. перипластидний простір, у якому знаходяться 80S рибосоми та унікальна ДНК-вмістна структура - нуклеоморф. Нуклеоморф відмежований від перипластидного простору власною двомембранною оболонкою (рис. 11.2).
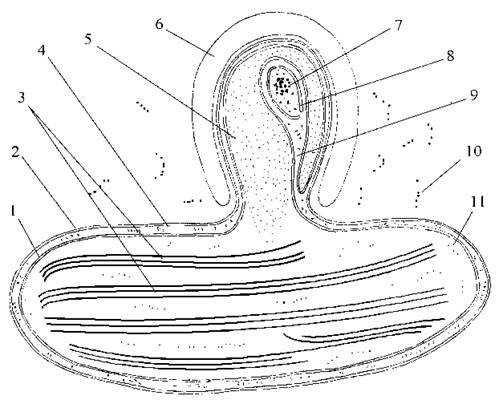
Рис. 11.2. Схема будови хлоропласту хлорарахніофітових водоростей: 1 - дві внутрішні мембрани оболонки хлоропласту; 2 - дві мембрани хлоропластної ендоплазматичної сітки; 3 - ламели, що складаються з трьох тилакоїдів; 4 - перипластидний простір з 80S рибосомами; 5 - піреноїд; 6 - парамілон; 7 - нуклеоморф; 8 - двомембранна оболонка нуклеоморфу; 9 — інвагінація хлоропластної ендоплазматичної сітки, в якій розташовується нуклеоморф; 10 - хлоропластні 70S рибосоми; 11 - цитоплазматичні 80S рибосоми (згідно до Hibberd, Norris, 1984).
Під хлоропластною оболонкою розташовуються тилакоїди, зібрані переважно по два або по три, рідше - поодинокі. Оперезуючий тилакоїд відсутній. В стромі хлоропласту також міститься хлоропластна ДНК, 70S рибосоми та великий, виступаючий за межі хлоропласту піреноїд. Цей піреноїд, на відміну від піреноїдів більшості інших водоростей, не пронизаний тилакоїдами. Навколо піреноїду, але завжди за межами хлоропласту, накопичується парамілон.
Мітохондріальний апарат представлений розгалуженим мітохондріоном з трубчастими кристами.
Джгутикові стадії у хлорарахніофітів представлені зооспорами з унікальним для водоростей планом будови: джгутик завжди один, субапікальний, спірально закручений навколо клітини і, на відміну від усіх інших водоростей, спрямований назад. Аксонема джгутика несе прості, дуже тонкі пірчасто розміщені мастигонеми. Стигма завжди відсутня.
Інші органели. У цитоплазмі є розвинений комплекс Гольджі, спостерігаються також травні вакуолі, які утворюються при захопленні ризоподіями дрібних одноклітинних водоростей, бактерій або часток детриту.
Розмноження та життєвий цикл
Розмноження відбувається поділом клітин надвоє або за допомогою зооспор. За несприятливих умов амебоїдні клітини можуть набувати сферичної форми, вкриватися оболонкою і переходити у кокоїдний стан.
У Chlorarachniophyta спостерігалось статеве розмноження, проте його особливості, місце проходження мейозу та зміна ядерних фаз лишаються не з’ясованими.
Життєвий цикл простий і являє собою цикломорфоз з чергуванням стадій поодиноких амебоїдних клітин, сітчастих колоній та зооспор. За несприятливих умов колонії розпадаються на окремі клітини, кожна клітина втягує ризоподії, набуває сферичної форми і перетворюється на спочиваючу кокоїдну акінету.
Система відділу
Всі відомі хлорарахніофітові належать до класу Chlorarachniophyceae, порядку Chlorarachniales, родини Chlorarachniaceae. Типовим представником є рід Chlorarachnion (див. рис. 11.1).
Походження хлорархніофітових та їх положення у системі евкаріот
На відміну від більшості евкаріот, хлорарахніофітові мають чотири геноми - ядерний, мітохондріальний, хлоропластний та геном нуклеоморфу. Походження першого та останнього геномів було досліджено молекулярно-біологічними методами на прикладі Chlorarachnion reptans.
Ядерний геном за результатами аналізу нуклеотидної послідовності гену, що кодує малу субодиницю РНК цитоплазматичних рибосом виявився спорідненим з геномами черепашкових амеб (Testacea), зокрема, з родів Euglypha та Paulinella (рис. 11.3). Геном нуклеоморфу виявися спорідненим з ядерними геномами зелених водоростей з порядку Volvocales.
На підставі цих результатів було запропоновано схему походження хлорарахніофітових, що добре узгоджується також із комплексом специфічних фенотипічних ознак представників відділу (рис. 11.4). Так, згідно з цією схемою, хлорарахніофітові виникають внаслідок ендосимбіозу амебоїдної гетеротрофної клітини, близько спорідненої з Testacea, та фотоавтотрофної зеленої водорості з Volvocales. Процес відбувається у два етапи.
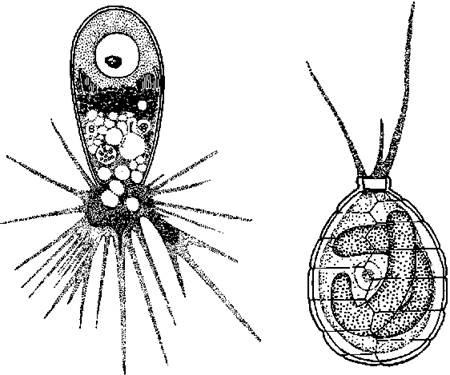
Рис. 11.3. Приклади найбільш споріднених з Chlorarachniophyta черепашкових амеб: Euglypha (ліворуч) та Paulinella (праворуч). Цікаво, що Paulinella у цитоплазмі містить симбіотичні синьозелені водорості - цианели (за Догель, 1981; Kies, 1974).
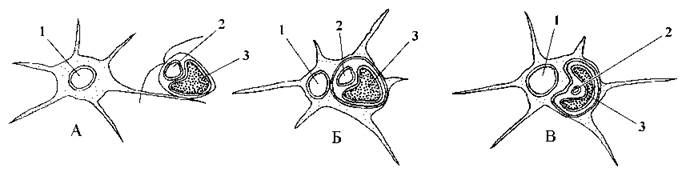
Рис. 11.4. Гіпотетична схема походження хлорарахніофітових водоростей: А - амебоїдна гетеротрофна клітина захоплює зелену водорость; Б - травна вакуоля з зеленою водоростю у цитоплазмі клітини-господаря; В - ендосимбіонт трансформується у хлоропласт, при цьому відбувається редукція його ядра. 1 - ядро амебоїдної клітини-господаря, 2 - ядро зеленої водорості, що трансформується у нуклеоморф, 3 - хлоропласт (за McFadden & al., 1997).
На першому етапі водорость захоплюється псевдоподіями амеби, і утворюється травна вакуоля. Це пояснює наявність чотирьох мембран навколо хлоропласту: дві внутрішні є мембранами хлоропласту зеленої водорості, третя - її плазмалемою, четверта - мембраною травної вакуолі амеби. Цитоплазма зеленої водорості, що містить 80S рибосоми, утворює перипластидний простір.
На другому етапі відбувається часткова редукція ядра зеленої водорості й перетворення його на нуклеоморф. Так, у Chlorarachnion reptans в нуклеоморфі міститься лише три хромосоми, що містять 145, 140 та 95 тис. пар нуклеотидних основ. Ці хромосоми зберігають гени, що кодують перипластидну рибосомальну РНК, та щонайменше десять генів, що кодують ферменти, які беруть участь у процесах реплікації, транскрипції та трансляції.
Таким чином, молекулярні та цитологічні дані свідчать, що Chlorarachniophyta належать до царства Тубулокристат, утворюючи разом з амебами-філозеями та деякими іншими амебоїдними організмами групу примітивних трубчасто-кристних організмів. Ця група отримала статус підцарства Амебо-флагеляти.