Альгологія - Костіков І.Ю. - 2009-2013
Розділ 10. Евгленофітові водорості – Euglenophyta
Відділ Euglenophyta нараховує біля 1000 видів мікроскопічних водоростей, поширених переважно у прісних континентальних водоймах. Невелика кількість видів евгленофітів відома з морських біотопів1. За палеоальгологічними даними, найдавніші викопні рештки Euglenophyta датуються віком біля 65 млн. років (кайнозой, третинний період).
Відділ включає не тільки фотоавтотрофні, але й значну кількість гетеротрофних видів. Окремі представники ведуть паразитичний спосіб життя. За винятком роду Colacium, всі евгленофітові водорості мають монадний тип будови.
Систематичні ознаки відділу
Пігменти та запасні поживні речовини
Для забарвленних представників характерна наявність хлорофілів а та b, β- та γ-каротинів. Основними ксантофілами є неоксантин та діадіноксантин. У незначних кількостях виявлено також діатоксантин, зеаксантин, ехіненон. Оскільки хлорофіли не маскуються ксантофілами, хлоропласти фотоавтотрофних евгленофітів забарвлені у зелений колір.
Основним продуктом асиміляції евгленофітових водоростей є парамілон. Він накопичується завжди поза межами хлоропласту у вигляді дрібних гранул або великих, добре помітних парамілій; парамілон може відкладатись також на виступаючих за межі хлоропласту поверхнях піреноїдів. З розчином йоду реакції не дає, від лугу набрякає. Додатковим асимілятом у Euglenophyta є масло. Окрім евгленофітів, парамілон виявлено лише у примнезіофітових водоростей з порядку Pavlovales та у хлорархніофітових водоростей.
Цитологічні ознаки
Клітинні покриви евгленофітових водоростей представлені пелікулою (рис. 10.1.). Пелікула утворена плазмалемою, під якою розташовуються поздовжні або спірально вигнуті білкові стрічки. У зоні стикання сусідніх стрічок утворюються виступи, які надають поверхні клітин характерну поздовжню або спіральну посмугованість. Під білковими стрічками пелікули розташовується сітка з мікротрубочок, яка з'єднується зі стрічками за допомогою зубців. З внутрішнього боку пелікули у багатьох видів Euglenophyta знаходяться заповнені слизом вакуолі - т.з. слизові тільця. При набряканні слизу вміст слизових тілець виходить через слизові канали на поверхню пелікули. За допомогою цього слизу клітини утворюють слизові капсули, слизові ніжки, приклеюються до субстрату.
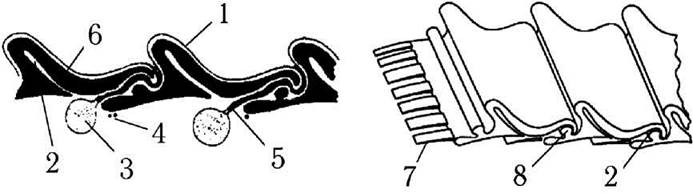
Рис. 10.1. Будова пелікули: 1 - плазмалема, 2 - внутрішня складка, 3 - слизове тільце, 4 - підстилаючі мікротрубочки, 5 - слизовий канал, 6 - білкова стрічка, 7 - з'єднувальний зубець пелікули, 8 - ребро (за Leedale, 1967).
Пелікула звичайно еластична, тому клітини здатні в процесі руху змінювати свою форму - витягуватись, стискатись і т.п. Клітини, які не зберігають у процесі руху постійну форму, називають метаболічними. У видів з неметаболічною формою клітин пелікула має жорстку конструкцію.
У деяких евгленофітових водоростей монадні клітини розміщуються всередині будиночків (наприклад, у видів роду Trachelomonas). Будиночки утворені слизовими тяжами, просякнутими солями марганцю та заліза. Будиночки мають отвір, з якого виходить джгутик монади. Як правило, навколо отвору з зовнішнього боку розміщується комірець.
Загальний план будови клітини. Клітини евгленофітових водоростей мають досить однотипну будову (рис. 10.2). На передньому кінці клітини знаходиться заглиблення - глотка, що складається з каналу та резервуару. Біля стінок резервуару розташовуються кілька пульсуючих вакуолей. Ці вакуолі по мірі заповнення рідиною зливаються між собою в одну велику вакуоль, яка виплескує свій вміст у резервуар. З дна глотки підіймаються два джгутики.
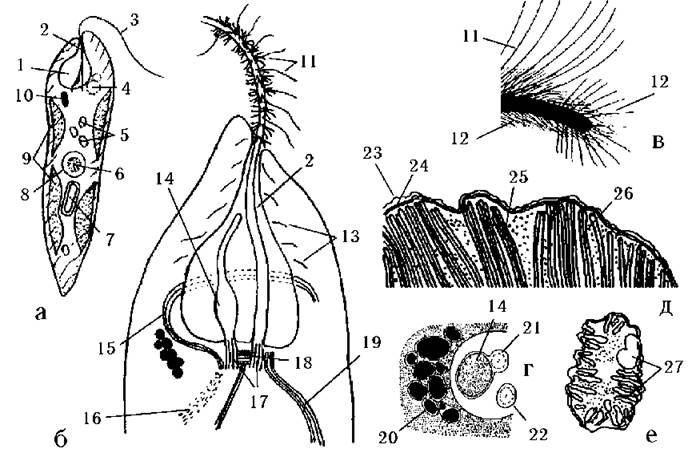
Рис. 10.2. Будова клітини евгленофітових водоростей: а - загальний вигляд клітини, б - будова глотки та коренева система джгутику, в - типи мастигонем, г - фоторецепторний апарат, д - фрагмент хлоропласту, е - зріз через мітохондріон. 1 - резервуар, 2 - канал глотки, 3 - джгутик, 4 - скоротлива вакуоля, 5 - парамілонові гранули, 6 - ендосома, 7 - парамілія, 8 - ядро, 9 - хлоропласти, 10 - стигма, 11 - однобічні довгі мастигонеми, 12 - апікальні та короткі проміжні мастигонеми, 13 - оперезуючі глотку мікротрубочки, 14 - парафлагелярне потовщення короткого джгутика, 15 - дорзальний мікротубулярний корінь, 16 - мікрофібрилярний корінь, 17 - базальні тіла джгутиків, 18 - поперечно-смугасте мікрофібрилярне з'єднання, 19 - вентральний мікротубулярний корінь, 20 - пігментна глобула вічка, 21 - аксонема короткого джгутика, 22 - аксонема довшого джгутика, 23 - зовнішня мембрана хлоропласту, 24 - внутрішні мембрани хлоропласту, 25 - тритилакоїдна ламела, 26 - двотилакоїдна ламела, 27 - дископодібні кристи (за Leedale, 1967; Асаул, 1986).
У частини гетеротрофних евгленофітових водоростей у цитоплазмі в районі глотки є органели для захоплення їжі - паличкоподібна органела або сифон. У цьому випадку часточки їжі, що потрапляють у глотку, захоплюються і перетравлюються у травних вакуолях. Неперетравлені рештки виштовхуються назовні на задньому кінці клітини. У видів родів Entosiphon та Peranemaвиявлені еджективні органели - трихоцисти, проте їх ультратонка будова та механізм відстрілювання практично не досліджені.
Ядерний апарат. Ядро евгленофітових водоростей має нетипову для еукаріот будову. Воно досить велике, добре помітне у живому стані навіть без застосування спеціальних барвників. Ядерна ДНК евгленофітів пов’язана з гістонами, але хромосоми завжди знаходяться у конденсованому стані. В центрі ядра розташовується велике ядерце, яке через здатність до самостійного поділу називають ендосомою. Ядерна оболонка при поділі ядра лишається інтактною, справжнє веретено поділу не утворюється, центриолі відсутні, а їх функції виконують базальні тіла джгутиків.
Фотосинтетичний апарат. Хлоропласти евгленофітових водоростей вкриті тримембранною оболонкою. Тилакоїди в хлоропластах евгленофітових водоростей групуються переважно по три. Інколи в хлоропастах спостерігаються голі напівзанурені піреноїди.
За результатами аналізу хлоропластної ДНК евгленофітів та за цитологічними даними вважають, що хлоропласти Euglenophyta виникли внаслідок симбіозу безбарвних евгленофітів з еукаріотичними зеленими водоростями. Тому дві внутрішні мембрани хлоропластів розглядають як власну хлоропластну оболонку зеленого пращура хлоропласту евгленофітів, а третю, зовнішню мембрану - або як залишок мембрани травної вакуолі евгленофтової клітини-господаря, або ж як залишок плазмалеми зеленої водорості - попередника хлоропласту Euglenophyta.
Мітохондрії. Звичайно клітина евгленофітових водоростей має одну, досить велику (до 10 мкм) розгалужену мітохондрію - т.з. мітохондріон. Будова мітохондріону евгленофітів унікальна серед водоростей, оскільки внутрішня мембрана утворює кристи специфічної дископодібної форми.
Джгутиковий апарат. Аксонеми джгутиків вкриті двома типами мастигонем: а) короткими, що з'єднуються у пучки по три-чотири і розташовуються на аксонемі спірально, і б) довгими поодинокими мастигонемами, які звичайно розташовуються гребінчасто. Довжина джгутиків звичайно неоднакова, у багатьох евгленофітів один з джгутиків редукований і не виходить за межі глотки; верхній кінець такого джгутика з'єднується з аксонемою довшого джгутика. Такі джгутики називають роздвоєними при основі. Інколи (наприклад, у евтрепцієвих водоростей) джгутики майже однакової довжини, або (у меноідієвих водоростей) джгутик один, при основі не роздвоєний (рис. 10.3). Від базальних тіл джгутиків відходять три мікротрубочкових корінця - дорзальний, вентральний та проміжний, які розташовуються навколо глотки і майже не заглиблюються в середню частину клітини (рис. 10.2).
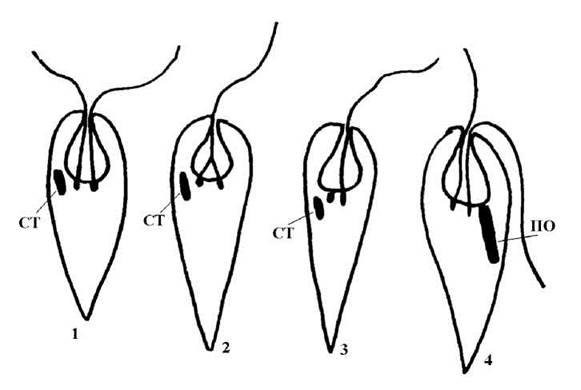
Рис. 10.3. Варіанти будови джгутикового апарату евгленофітових водоростей: 1 - джгутики майже однакові, 2 - джгутик один, роздвоєний при основі, 3 - джгутик один, не роздвоєний при основі, 4 - гетероморфні та гетеродинамічні джгутики. СТ - стигма, ПО - паличкоподібна органела.
При основі одного з джгутиків (рідше - обох) знаходиться потовщення - парафлагелярне тіло, що виконує фоторецепторні функції. Біля парафлагелярного тіла у багатьох Euglenophytaрозміщується стигма. Стигма побудована з кількох шарів пігментних глобул, кожна глобула відокремлена від цитоплазми власною мембраною. Стигма евгленофітів завжди розташована поза межами хлоропласта і виконує функцію ширми, яка регулює кількість світла, що потрапляє на фоторецептор - парафлагелярне тіло.
У деяких видів джгутики відсутні, хоча зберігаються їх базальні тіла. Такі клітини рухаються червеподібно, за рахунок метаболічних вигинів клітини.
Розмноження та життєві цикли
Розмноження евгленофітових водоростей відбувається шляхом поділу навпіл у рухливому або нерухомому (нерідко - пальмелевидному) стані. При поділі у рухливому стані в клітині утворюється новий джгутиковий апарат, далі ядро з центральної частини клітини мігрує вперед, до резервуару глотки. Потім всередині ядерної оболонки утворюється веретено поділу, причому базальні тіла джгутиків виконують функції центриолей. Далі відбувається поділ ендосоми та ядра, і за рахунок утворення апікальної борозни - цитокінез (рис. 10.4).
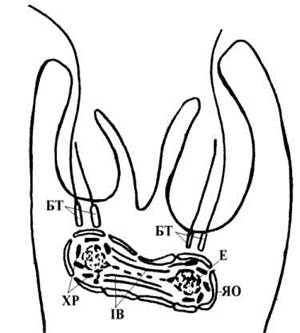
Рис. 10.4. Схема поділу клітини евгленофітових водоростей (телофаза). БТ - базальні тіла джгутиків, ЯО - ядерна оболонка, ХР - хромосоми, Е - ендосома, ІВ - закрите інтерзональне веретено поділу.
Проте більш поширеним є поділ у нерухомому стані. В цьому випадку клітина зупиняється та скидає джгутик. Якщо поділ відбувається у пальмелевидному стані, то слизові тільця виділяють слиз, а клітина набуває більш - менш сферичної форми. Далі відбувається реплікація базальних тіл джгутиків, утворення закритого веретена поділу, каріокінез та цитокінез (рис. 10.5).
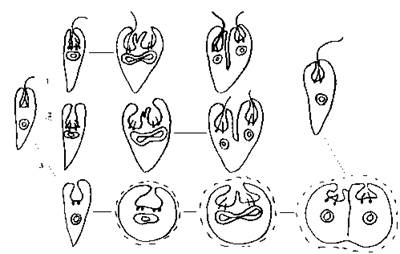
Рис. 10.5. Типи поділу клітин у евгленофітових водоростей: 1 - поділ у рухливому стані, 2 - поділ у нерухомому стані, 3 - поділ у пальмелевидному стані.
У трьох видів (Scytomonas subtilis, Phacus pyrum, Euglena sp.) описаний хологамний та автогамний статевий процес з зиготичною редукцією. Проте в цілому статеве розмноження для Euglenophyta не характерне.
Життєві цикли евгленофітових водоростей досить прості, здебільшого вони відбуваються без чергування гаплоїдної та диплоїдної ядерних фаз, тобто представлені цикломорфозом. Найчастіше цикломорфоз евгленофітів полягає у зміні монадного та пальмелевидного станів. Пальмелевидний стан спостерігається звичайно у несприятливих умовах.
Особливості живлення
Живлення різних представників Euglenophyta може відбуватись за рахунок фотосинтезу та осмотрофного або голозойного споживання органічних речовин. Фотоавтотрофні види здатні рости на мінеральних поживних середовищах, проте найкращий ріст спостерігається при додаванні у середовище органічних сполук. У темряві на органічних середовищах клітини деяких видів можуть втрачати хлорофіл, накопичувати феофітин, зберігаючи при цьому здатність до росту та розмноження. В знебарвлених клітинах замість хлоропластів спостерігаються лейкопласти. При перенесенні таких культур на світло клітини відновлюють хлорофіл, і лейкопласти перетворюються на хлоропласти. Знебарвлені культури можна отримати і в умовах освітлення - дією антибіотиків, екстремальних температур, ультрафіолетовим опромінюванням.
Паразитичні безбарвні водорості з порядку Euglenamorphales при перенесенні на штучні мінеральні середовища на світлі синтезують хлорофіл і переходять на фотоавтотрофне живлення.
Вільноіснуючі безбарвні Euglenophyta є облігатними гетеротрофами. Серед цієї групи частина видів живиться лише осмотрофно, за морфологією нагадує знебарвлених фотоавтотрофних представників. Друга частина видів живиться осмотрофно та голозойно, не має морфологічних "двійників" серед забарвлених форм, у багатьох є паличкоподібна органела або сифон.
Система відділу
Відділ включає один клас - Euglenophyceae, та три порядки - Euglenales, Peranematales, Euglenamorphales. В основу поділу на порядки покладено тип живлення та ступінь спорідненості безбарвних форм з забарвленими (рис. 10.6).
1. Порядок Euglenales. Об’єднує всіх забарвлених представників та безбарвні види, які вторинно втратили хлоропласти. Останні живляться осмотрофно і за морфологією нагадують забарвлені форми. В межах порядку є види з двома джгутиками, які виходять за межі глотки (наприклад, забарвлена Eutreptia та безбарвна Distigma - родина Eutreptiaceae), з одним не роздвоєним при основі джгутиком (родина Menoidiaceae; всі представники цієї групи безбарвні, основними родами є Menoidium та Gyropaigne), та види з одним джгутиком, що виходить за межі глотки та роздвоєний при основі. Ця група (родина Euglenaceae) є найбільш поширеною та багатою на види і займає провідне місце у системі Euglenophyta в цілому. Основними родами є Euglena (об’єднує забарвлені види з несплощеними, слабо метаболічними клітинами), Trachelomonas (клітини забарвлені та знаходяться в будиночках), Phacus (клітини забарвлені та сильно стиснуті з боків), Astasia (в морфологічному відношенні нагадує евглену, проте позбавлена хлоропластів та стигми) (рис. 10.7).
2. Порядок Peranematales. Включає лише безбарвні види, які не мають забарвлених аналогів. Живляться перанематальні водорості осмотрофно та голозойно, багато представників мають пристосування для активного захоплення їжі - паличкоподібну органелу або сифон. До порядку входять види як з одним нероздвоєним при основі джгутиком, так і з двома гетеродинамічними джгутиками. В останньому випадку рух відбувається за допомогою переднього джгутика, аксонема якого звичайно рухлива тільки на кінці; другий джгутик виконує функції стерна. З видів, позбавлених паличкоподібної органели (родина Petalomonadaceae) найбільш поширеними є види роду Sphenomonas, з групи видів з паличкоподібними органелами (родина Peranemataceae) - Peranema та Heteronema.
3. Порядок Euglenamorphales включає безбарвні види, що паразитують у тілі водних тварин. При вилученні з тіла тварини-господаря та перенесенні на світло клітини евгленоморфальних водоростей синтезують хлорофіл та набувають зеленого забарвлення. Типовим представником цього порядку є Euglenamorpha.
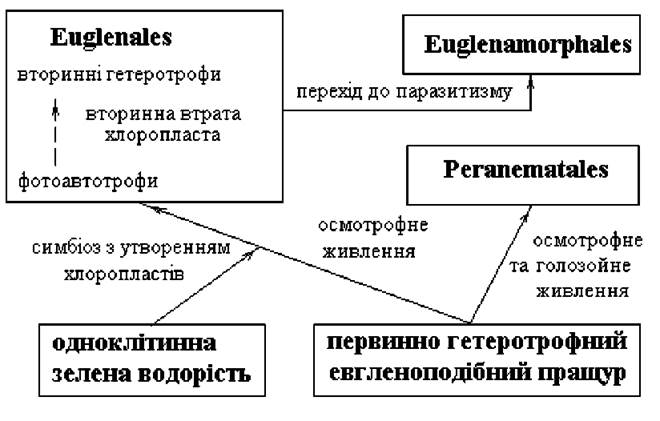
Рис. 10.6. Схема родинних зв’язків різних порядків Euglenophyta.
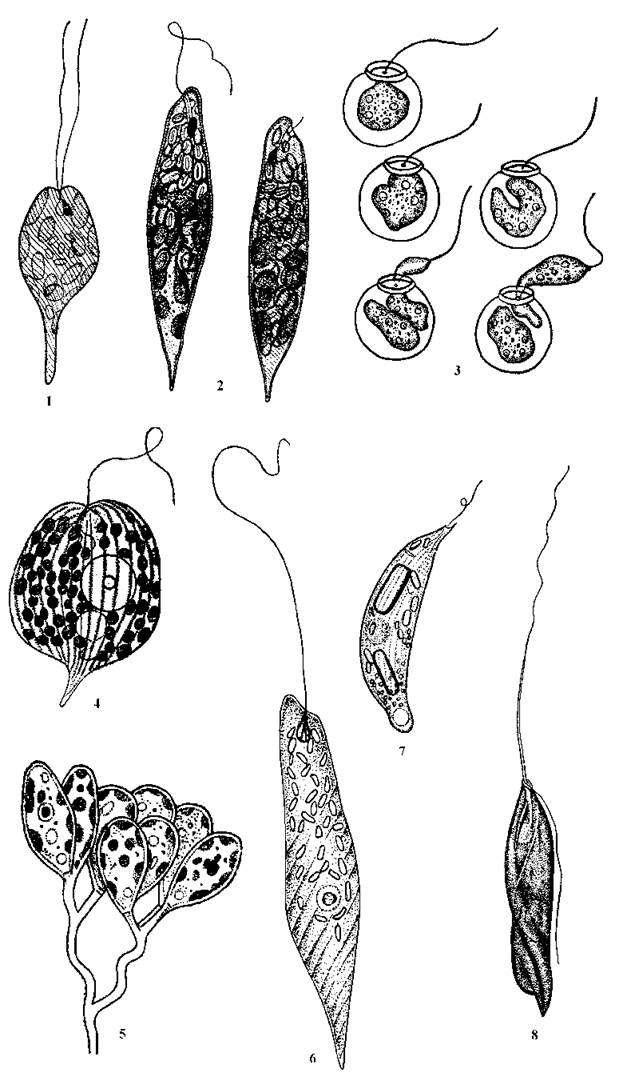
Рис. 10.7. Характерні представники відділу Euglenophyta: 1 - Eutreptia, 2 - Euglena, 3 - Trachelomonas (монади у будиночках та розмноження), 4 - Phacus, 5 - Colacium, 6 - Astasia, 7 - Menoidium, 8 - Heteronema(за Асаул, 1975, 1986).
Поширення, екологія та значення
Переважна більшість евгленофітових водоростей мешкає у прісних континентальних водоймах. Найбільш інтенсивно види Euglenophyta розвиваються в ефемерних водоймах, ставках, річках з уповільненою течією, болотах тощо. Значно бідніше відділ представлено в озерах, водосховищах та лиманах. Деякі евгленофіти (наприклад, Euglena proxima, Trachelomonas volvocina) знайдені у позаводних умовах на вологому ґрунті.
Більшість видів Euglenophyta теплолюбні, їх температурний оптимум знаходиться у межах 20...27 °С, хоча відомі й евритермні види, здатні розвиватись в діапазоні температур від 0 до 30 °С (деякі Trachelomonas, Euglena, Phacus, Astasia). Відносно активної реакції середовища евгленофіти є слабкими ацидофілами (оптимум pH становить 5.5-7.0). У лужному середовищі (pH≥8) кількість видів різко зменшується. Це пов’язують, у першу чергу, зі зменшенням у воді концентрації розчинених солей заліза, необхідних для нормальної життєдіяльності представників цього відділу. Евгленофітові водорості дуже чутливі до ступеню мінералізації води і, як правило, при концентрації солей вище 5 г/л гинуть. Винятком є безджгутикова Euglena vermicularis, яка нормально вегетує навіть у гіпергалійних озерах, при солоності до 100 г/л. Наявність розчинених у воді органічних сполук стимулює розвиток Euglenophyta (особливо - гетеротрофних), значна кількість видів інтенсивно розвивається у водоймах очисних споруд. У невеликих водоймах, багатих на органіку (найчастіше - у калюжах), евгленофітові водорості здатні викликати локальні "цвітіння" води, які, на відміну від "цвітіння" синьозеленими водоростями, не токсичні.
У практичній діяльності людини евгленофітові водорості використовуються, перш за все, як надійні біоіндикатори для оцінки якості води та ступеню її забруднення. Крім того, представники Euglenophyta (особливо Euglena viridis) є модельними об'єктами у біохімічних, біофізичних, фізіологічних та генетичних дослідженнях.
Положення Euglenophyta в ботанічних та зоологічних системах
Положення евгленофітових у системі живих істот довгий час лишалось невизначеним і було предметом численних дискусій. Перші відомості про евгленофітів були отримані протозоологами і опубліковані в зоологічних виданнях. Починаючи з 20-х років питаннями систематики, поширення, фізіології, практичного використання цієї групи займаються майже виключно альгологи. Зоологи звертаються до Euglenophyta лише у зв’язку з питаннями класифікації Protozoa, при цьому евгленофіти до кінця 70-х років наполегливо вміщуються до класу Mastigophora як ряд Euglenida. У пізніших системах протозоологів (див. наприклад, Серавин, 1980; Patterson, 1994) таксономічний ранг всіх фітофлагелят підвищений і до початку - середини 90-х років евгленофіти наводяться як клас Euglenomonada в межах типу Sarcomastigophora або як самостійний клас Euglenida в межах типу Euglenozoa (другим класом в цьому типі є Kinetoplastida).
У ботанічних системах евгленофітові водорості з 20-х років розглядають як окремий відділ або як самостійний клас в системі зелених водоростей. Остання точка зору недостатньо переконлива, оскільки Euglenophyta подібні до зелених водоростей лише за складом пігментів.
За сучасними цитологічними, і особливо - молекулярно-біологічними даними Euglenophyta є одним з найбільш давніх відділів еукаріот, що разом з Kinetoplastida розташовується при основі еукаріотного філогенетичного дерева. Первинні евгленофітові водорості були облігатними гетеротрофами з тваринною стратегією життя. Фотоавтотрофні евгленофіти є більш молодою групою, яка з’явилась внаслідок ендосимбіозу гетеротрофної клітини-господаря з одноклітинною зеленою водоростю. Остання була далі трансформована у хлоропласт Euglenophyta. За своєрідними мітохондріями з дископодібними кристами та за нуклеотидними послідовностями деяких ядерних генів (зокрема, гену, що кодує цитоплазматичну рибосомальну РНК), евгленофіти та кінетопластиди є близькими до схізопіренід та акразієвих слизовиків. На підставі цієї схожості ці чотири групи деякі дослідники навіть вважають самостійним царством Дискокристат.
В цілому, на прикладі Euglenophyta можна бачити, як в межах однієї систематичної групи йшло випробування рослинної та тваринної стратегій життя. Питання про те, чи є евгленофіти/евгленозої рослинами чи тваринами позбавлено сенсу, оскільки прірва між евгленою та будь-якою багатоклітинною твариною або вищою рослиною набагато більша, ніж філогенетична відстань, цитологічна та генетична різниця між, наприклад, собакою та пшеницею.
1 Вважають, що морські евгленофіти, що мешкають у прибережних водах - це численна та різноманітна, проте майже не досліджена група водоростей. Відомі випадки, коли евгленофітові водорості викликали морське "цвітіння" води біля узбережжя Норвегії.