Альгологія - Костіков І.Ю. - 2009-2013
Розділ 9. Синьозелені водорості – Cyanophyta
Синьозелені водорості (Cyanophyta), або цианопрокаріоти (Cyanoprocaryota), або цианобактерії (Cyanobacteria) є єдиним відділом прокаріотичних організмів, що здатні до оксигенного фотосинтезу. Нараховує біля 2000 видів. Синьозелені водорості бувають одно- та багатоклітинні, переважно - мікроскопічні, лише деякі колоніальні види сягають значних розмірів (наприклад, Nostoc commune або види Gloeotrichia).
Синьозелені водорості є однією з найдавніших груп на планеті. Вважають, що Cyanophyta виникли біля 3.5-3.8 млрд. років тому (для порівняння - вік Землі за даними аналізу метеоритного свинцю складає 4.6 млрд. років, перші еукаріоти з’явились біля 1.5 млрд. років тому).
Найдавніші викопні рештки синьозелених водоростей (т.з. строматоліти) датуються віком біля 3.2 млрд. років. З цього часу і протягом майже 2 млрд. років Cyanophyta панували на Земній кулі. Завдяки здатності синьозелених водоростей до оксигенного фотосинтезу на планеті виникла киснева атмосфера.
Систематичні ознаки відділу
Пігменти та запасні поживні речовини
Синьозелені водорості забарвлені переважно у блакитно-зелений колір, в екстремальних умовах частіше мають забарвлення з різними відтінками червоного кольору. Відомо також кілька представників, у яких клітини зелені.
До недавнього часу вважалося, що склад пігментів у Cyanophyta досить одноманітний: зеленого відтінку клітинам надає хлорофіл "а". Червоний або блакитний колір обумовлені наявністю значної кількості фікобілінових пігментів - фікоцианіну, аллофікоцианіну (сині пігменти) та фікоеритрину (червоний пігмент). Каротиноїди представлені лише β-каротином, ксантофілами лютеінового циклу (лютеіном та зеаксантином) та специфічними ксантофілами синьозелених водоростей - переважно осцилоксантином, міксоксантином, афаніцином та афанізофілом.
До Cyanophyta належить також невелика, проте цікава група зелених прокаріотичних водоростей, відкрита у 1976 р. - т.з. прохлорофітові водорості. Ці водорості, крім хлорофілу "а" мають також хлорофіл "b" або хлорофіл-подібний пігмент (дивініл-хлорофіл а-подібний Mg-порфірин), інколи також α-каротин.
Майже у всіх Cyanophyta основним продуктом асиміляції є глікогеноподібний полісахарид - крохмаль синьозелених водоростей, зрідка - полісахарид, що нагадує справжній рослинний крохмаль (у "прохлорофітів"). Крім вуглеводів, більшість синьозелених водоростей запасає також цианофіцин та волютин. Цианофіцин є полімером амінокислот аргініну та аспарагіну і використовується як резервне джерело доступного для клітини азоту. Волютин є запасною речовиною фосфорних сполук і утворює т.з. поліфосфатні гранули.
Цитологічні ознаки
Клітини Cyanophyta мають типову прокаріотичну будову (рис. 9.1): вони позбавлені морфологічно оформленого ядра, мітохндрій, хлоропластів, ендоплазматичної сітки, комплексу Гольджі, лізосом. У Cyanophyta відсутні будь-які структури, побудовані з мікротрубочок - джгутики, їх базальні тіла, мікротубулярні елементи цитоскелету, центриолі, веретено поділу. Синьозелені водорості не здатні до мітозу, мейозу та типового для еукаріот статевого процесу.
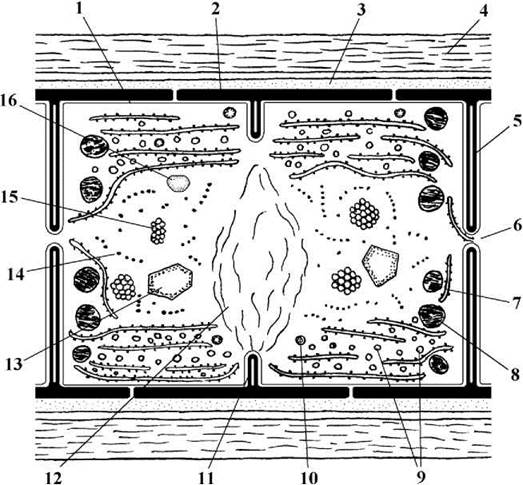
Рис. 9.1. Будова клітини синьозеленої водорості: 1 - плазмалема; 2, 3, 3a, 3b - клітинна оболонка: муреїновий шар (2); периплазматичний простір з геміцелюлозно-пектиновими фібрилами (3); зовнішня мембрана (outer membrane) (3а); зовнішній зубчастий шар з осциліновими фібрилами (3b); з’єднувальний поровий комплекс (3с); 4 - слизова піхва; 5 - поперечна перегородка; 6 - пора з плазмодесмою; 7 - тилакоїд з фікобілісомами; 8 - гранула цианофіцину; 9 - гранули крохмалю синьозелених водоростей; 10 - ліпідна глобула; 11 - зачаткова поперечна перегородка; 12 - нуклеоїд; 13 - поліедральне тіло; 14 - полісома; 15 - газова вакуоля; 16 - поліфосфатне тіло (схематизовано за Кондратьева, 1989; Anagnostidis, Komarek, 1986; 1988; 1990; Hoiczyk, Baumeister, 1998).
Клітинні покриви у Cyanophyta - це клітинна оболонка. Вона розташовується назовні від плазмалеми і складається з кількох шарів: а) шару муреіну; б) периплазматичного простору, виповненого переважно пектином та геміцелюлозою; в) зовнішньої мембрани; г) зубчастого шару з поверхневими фібрилами глікопротеїну осциліну. Через муреїновий шар та периплазматичний простір біля поперечної перегородки під кутом 30-40° до поздовжньої вісі клітини можуть проходити т.зв. з’єднувальні порові комплекси, через які назовні секретується локомоторний слиз. Секреція цього слизу обумовлює здатність багатьох синьозелених водоростей до активного ковзаючого або обертального руху. В цілому, клітинні оболонки Cyanophyta подібні до оболонок грамнегативних бактерій, але вміст муреїну по відношенню до загальної маси оболонки вищий, ніж у бактерій (22-52% замість 5-10%).
Назовні від клітинної оболонки у більшості видів виділяються пектинові речовини, що утворюють колоніальний слиз або слизові піхви.
Ядерний апарат. Справжнього ядра у синьозелених водоростей немає, а його функції виконує т.з. нуклеоїд. Нуклеоїд - це одна або кілька молекул ДНК, замкнених у кільце, що розташовується безпосередньо у цитоплазмі. ДНК нуклеоїду не пов’язана з білками-гістонами і не має хромосомної організації, хоча і розглядається як аналог однієї хромосоми. Нуклеоїд прикріплюється безпосередньо до плазмалеми. Звичайно основна маса нуклеоїду зосереджується у центрі клітини, через що цю зону цитоплазми називають нуклеоплазмою або центроплазмою.
Перед початком поділу ДНК нуклеоїду реплікується, далі точки прикріплення материнської та дочірньої молекул ДНК до плазмалеми починають розходитися, і в проміжку між ними утворюється інвагінація плазмалеми, яка і розділяє остаточно ядерні апарати двох нових клітин (рис. 9.2).
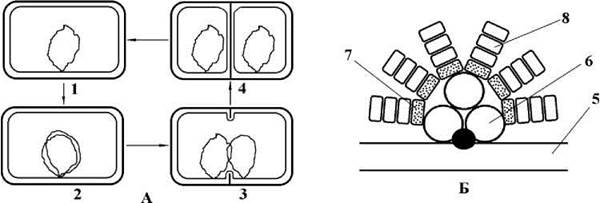
Рис. 9.2. Синьозелені водорості. А - схема поділу нуклеоїду (1 - клітина перед початком поділу; 2 - реплікація нуклеоїду; 3 - початок розходження точок прикріплення нуклеоїду до плазмалеми; 4 - дочірні клітини). Б - будова фікобілісоми (5 - мембрана тилакоїду; 6 - аллофікоцианін; 7 - фікоцианін; 8 - фікоеритрин) (Б - за Gantt, 1981).
Крім нуклеоїду, у багатьох синьозелених водоростей у цитоплазмі наявні плазміди - невеликі молекули ДНК, що також замкнені у кільце, проте, на відміну від нуклеоїду, вільно розташовуються у цитоплазмі.
Обмін генетичною інформацією між різними клітинами синьозелених водоростей здійснюється парасексуально - шляхом трансформації та кон’югації. При трансформації ДНК клітини-донора у розчиненому стані потрапляє у зовнішнє середовище і звідти - до клітини-рецепієнта, "вбудовуючись" у її нуклеоїд. При кон’югації дві клітини безпосередньо з'єднуються вузьким цитоплазматичним каналом, по якому і здійснюється обмін молекулами ДНК, їх ділянками або плазмідами.
Фотосинтетичний апарат представлений тилакоїдами, що є похідними інвагінацій плазмалеми. Тилакоїди не відокремлюються від цитоплазми двомембранною оболонкою, тобто фотосинтетичний апарат не організований у вигляді пластид. Тилакоїди звичайно розташовуються поодиноко, зрідка (у деяких "прохлорофітів") зібрані у стопки, які дещо нагадують грани. Звичайно тилакоїди розташовуються у периферичному шарі цитоплазми. В оптичний мікроскоп зона розташування тилакоїдів інтенсивно забарвлена, через що її називають хроматоплазмою. В мембрани тилакоїдів "вбудовані" молекули хлорофілу та додаткові пігменти - каротини та ксантофіли.
На поверхні тилакоїдів у всіх представників, за винятком "прохлорофітів" розташовуються також особливі "антенні" структури - фікобілісоми. Кожна фікобілісома утворена трьома глобулами аллофікоцианіну, що безпосередньо закріплюються у мембрані тилакоїду. Від глобул аллофікоцианіну відходять шість "променів", що при основі складаються з глобул фікоцианіну, а на верхівці - з фікоеритрину (рис. 9.2). Кількість глобул фікоеритрину може змінюватись залежно від спектральної характеристики світла: при переважанні короткохвильового синього та фіолетового опромінювання їх кількість збільшується, і клітини набувають червоного забарвлення, і навпаки, при фотосинтезі у середній да довгохвильовій частинах спектру кількість фікоеритрину зменшується, і клітини набувають блакитно-зеленого забарвлення. Це явище отримало назву хроматичної адаптації.
До елементів фотосинтетичного апарату у Cyanophyta опосередковано відносять також поліедральні тіла. Вони звичайно розташовуються у хроматоплазмі і мають вигляд кутастих включень. Поліедральні тіла утворені ферментом рибульозо-дифосфат-карбоксилазою (RuBisCo), який бере участь в асиміляції вуглекислого газу у темновій фазі фотосинтезу. Поліедральні тіла є попередником піреноїду евкаріотичних водоростей.
Інші цитоплазматичні структури. Всі Cyanophyta мають рибосоми, дифузно розміщені у цитоплазмі. Ці рибосоми дрібніші, ніж цитоплазматичні рибосоми евкаріот (їх розмір становить 21 х 29 нм, коефіцієнт седиментації - біля 70S, замість 22 х 32 нм та 80S, як це має місце у цитоплазмі евкаріотичних клітин). Рибосоми, подібні до Cyanophyta, виявлені у хлоропластах та мітохондріях евкаріот.
Включення представлені гранулами запасних поживних речовин, а також газовими вакуолями. Гранули резервних полісахаридів (крохмалю синьозелених водоростей, крохмалеподібного полісахариду "прохлорофітів") дрібні, у звичайних вегетативних клітин вони помітні лише в електронний мікроскоп, і локалізуються серед тилакоїдів. У спочиваючих клітинах - акінетах, комплекси, утворені полісахаридними гранулами можна спостерігати і на світло-оптичному рівні.
Цианофіцинові та поліфосфатні гранули великі - до 2 мкм у діаметрі. Перші у багатьох нитчастих синьозелених водоростей концентруються біля поперечних клітинних перегородок, другі більш-менш дифузно розсіяні по цитоплазмі.
У деяких (переважно - планктонних) видів у цитолазмі є газові вакуолі. В оптичний мікроскоп вони мають вигляд невеликих ділянок цитоплазми, що сильно заломлюють світло. За допомогою електронного мікроскопу встановлено, що такі ділянки складаються з багатьох шестикутних щільно притиснутих одна до одної комірок, кожна з яких відмежовується від інших власною оболонкою, яка являє собою не фосфоліпідну, а білкову мембрану. Комірки при сприятливих умовах заповнюються газом, і зменшують питому вагу клітини. При несприятливих умовах газ дифундує, газові вакуолі зменшуються в об’ємі та злипаються (колапсують), питома вага клітини збільшується і водорость осідає на дно, де "очікує" покращення умов для фотосинтезу.
Фізіолого-біохімічні особливості відділу
Серед прокаріот Cyanophyta є єдиним відділом, представники якого мають водночас першу та другу фотосистеми, через що здатні до оксигенного фотосинтезу. Цей процес здійснюється за універсальним для рослин рівнянням:
2H2O + CO2 + hγ = [CH2O] + H2O + O2
В той же час вони мають принаймні дві важливі фізіолого-біохімічні особливості, які можна розглядати як фізіологічні атавізми: це здатність здійснювати аноксигенний фотосинтез та фіксацію атмосферного азоту.
Аноксигенний фотосинтез у Cyanophyta спостерігається зазвичай в анаеробних умовах при наявності у середовищі достатньої кількості сірководню. В цьому випадку як донор протонів та електронів для здійснення фотосинтезу синьозелені водорості використовують сірководень:
2H2S + CO2 + hγ = [CH2O] + H2O+ 2S
Найчастіше кінцевим продуктом окиснення сірководню є молекулярна сірка, що здатна накопичуватися у цитоплазмі. Проте деякі морські Cyanophyta окиснюють сірководень до сульфідів. Подібний шлях метаболізму є основним для багатьох фотосинтезуючих бактерій-грацилікутів, проте у Cyanophyta цей шлях є другорядним.
Фіксація атмосферного азоту (азотфіксація) вважається одним з найдавніших фізіологічних процесів, який з’явився ще у архебактерій. В той же час жодний евкаротичний організм до азотфіксації не здатний. При азотфіксації молекулярний азот відновлюється до сполук амонію, і в цій формі включається в основні шляхи метаболізму клітини. Процес азотфіксації каталізується нітрогеназним ферментним комплексом, який повністю інгібується молекулярним киснем. Тому азотфіксація здійснюється лише в анаеробному середовищі.
Сьогодні здатність до фіксації азоту в анаеробних умовах виявлена у досить широкого кола одно- та багатоклітинних синьозелених водоростей. Проте серед багатоклітинних Cyanophyta є чимало видів, що фіксують атмосферний азот також у присутності кисню. У цієї групи нітрогеназний комплекс локалізований в особливих клітинах - гетероцистах, які мають кілька пристосувань, що запобігають проникненню вільного кисню у клітину, і тим самим на фоні аеробного зовнішнього середовища створюють у цитоплазмі клітини-гетероцисти анаеробні умови.
Будова талому
Синьозелені водорості бувають як одно-, так і багатоклітинними. Як одноклітинні, так і багатоклітинні представники можуть бути поодинокими або утворювати різноманітні колонії, в яких індивіди утримуються разом найчастіше за допомогою колоніального слизу.
Основним і обов'язковим елементом тіла багатоклітинних водоростей є трихом. Трихом являє собою сукупність фізіологічно пов'язаних клітин. Зв'язок відбувається за допомогою плазмодесм, що проходять через пори поперечних перегородок клітин. Назовні від трихому можуть знаходитись структуровані слизові утвори - піхви, які звичайно відіграють захисну функцію. Трихом разом із піхвою називають ниткою. У видів, позбавлених піхв, трихом і нитка є синонімічними поняттями.
Трихоми можуть бути нерозгалуженими та розгалуженими. Клітини нерозгалужених трихомів діляться лише в одній площині. Якщо клітини здатні до поділу у кількох площинах, трихоми галузяться. Цей тип галуження називають справжнім. Коли ж галузяться нитки, а самі трихоми лишаються нерозгалуженими, то галуження вважають несправжнім (рис. 9.3).
Основні типи клітин
У синьозелених водоростей розрізняють три основні типи клітин: а) вегетативні клітини, що здійснюють фотосинтез та здатні до поділу; б) гетероцисти - спеціалізовані клітини, що виконують функцію фіксації атмосферного азоту; в) акінети - спочиваючі клітини, за допомогою яких водорості переносять несприятливі умови (рис. 9.3). Вегетативні клітини властиві всім Cyanophyta, гетероцисти та акінети - лише частині багатоклітинних представників. Трихоми, що складаються лише з вегетативних клітин, називають гомоцитними, а ті, що складаються з вегетативних клітин, гетероцист та акінет - гетероцитними.
Гетероцисти містять нітрогеназний ферментний комплекс і виконують функції фіксації атмосферного азоту. Цією функцією, зокрема - необхідністю захищати нітрогеназу від згубної дії кисню - обумовлені морфологічні особливості гетероцист: вони безбарвні, нездатні до фотосинтезу, мають потовщену, звичайно - потрійну оболонку. Необхідні органічні речовини гетероцисти отримують від сусідніх вегетативних клітин. Зв'язок з останніми здійснюється за допомогою плазмодесм, що проходять крізь пори у поперечній перегородці клітин. З боку гетероцисти пори закриті добре помітною в оптичний мікроскоп пробкою, що не пропускає у гетероцисту кисень, проте є проникною для органічних речовин та води.
Акінети утворюються з вегетативних клітин і також мають потовщену оболонку. Проте вони зберігають хлорофіл та накопичують велику кількість запасних поживних речовин, звичайно - глікогеноподібного полісахариду.
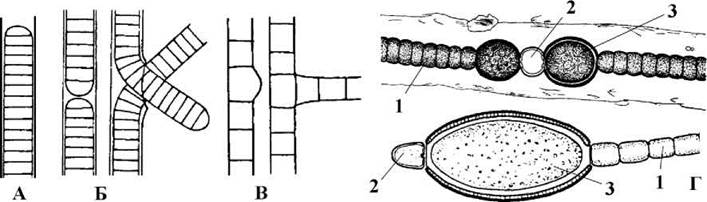
Рис. 9.3. Галуження ниток (А-В) та типи клітин (Г) у багатоклітинних Cyanophyta. А - нерозгалужені нитки, Б - нитки з несправжнім галуженням, В - справжнє галуження; Г - нитки з гетероцитними трихомами: 1 - вегетативні клітини, 2 - гетероцисти, 3 - акінети (за Anagnostidis, Komarek, 1988; Кондратьєва, 1968)
Поділ клітин та розмноження
В основі процесів розмноження у всіх синьозелених водоростей лежить клітинний поділ. При клітинному поділі спочатку плазмалема, а за нею - і муреїновий шар клітинної оболонки доцентрово вростають у протопласт, розділяючи клітину на дві (зрідка - більше) рівні або нерівні частини. Клітинний поділ у синьозелених водоростей може бути повним або неповним.
При повному поділі плазмалема та клітинна перегородка повністю відокремлюють дочірні клітини одна від одної. При цьому пори та плазмодесми не утворюються, і кожна дочірня клітина представляє собою фізіологічно самостійний індивід. Повний поділ властивий одноклітинним Cyanophyta.
Повний поділ поділяють на три типи: нормальний (бінарний), рівний багаторазовий та нерівний (рис. 9.4).
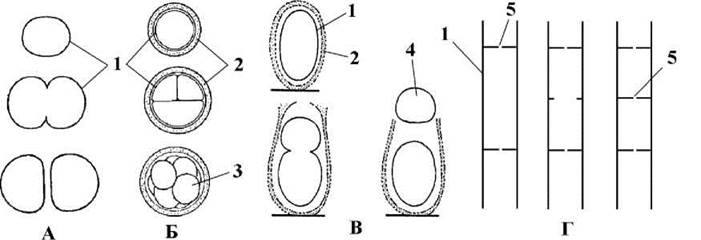
Рис. 9.4. Типи поділу у синьозелених водоростей: А - нормальний (бінарний); Б - рівний багаторазовий з утворенням наноцитів; В - нерівний з утворенням екзоспор; Г - неповний поділ. 1 - клітинна оболонка, 2 - слизова обгортка, 3 - наноцит, 4 - екзоцит, 5 - пора (за Komarek, Anagnostidis, 1986; Anagnostidis, Komarek, 1988)
При нормальному поділі материнська клітина ділиться на дві однакові за розміром дочірні клітини, що звичайно більш-менш відокремлюються одна від одної. Цей тип поділу притаманний майже всім одноклітинним синьозеленим водоростям.
При рівному багаторазовому поділі відбувається кілька швидких послідовних поділів навпіл, і дочірні клітини деякий час утримуються разом слизовою обгорткою материнської клітини. Під час поділу вони не ростуть, і за розмірами стають значно меншими, ніж материнська клітина. Тому їх називають наноцитами1.
Нерівний поділ дещо нагадує брунькування: материнська клітина ділиться на дві неоднакові за розміром клітини. Зрідка ці клітини деякий час лишаються з'єднаними слизовою обгорткою материнської клітини, проте частіше по закінченні поділу слизова обгортка з боку меншої клітини руйнується, і остання відокремлюється від більшої клітини. Меншу клітину в подібному випадкуназивають екзоцитом2.
Неповний бінарний поділ в принципі подібний до нормального бінарного, проте поперечна перегородка не повністю розділяє дочірні клітини, і вони лишаються з'єднаними плазмодесмою, що проходить через пору в поперечній клітинній перегородці. Внаслідок такого поділу утворюються багатоклітинні ниткоподібні трихоми.
Короткі рухливі фрагменти трихомів, що складаються з 2-50 клітин, називають гормогоніями, нерухомі - гормоцитами. Гормогонії та гормоцити є спеціалізованими репродуктивними структурами багатоклітинних Cyanophyta.
Колоніальні одно- та багатоклітинні синьозелені водорості здатні розмножуватись також шляхом фрагментації колоній.
Система відділу
Сьогодні існує кілька систем Cyanophyta, які в принципі можна умовно поділити на три групи: класичні морфологічні, морфолого-цитологічні та
молекулярно-біологічні. Ці три групи систем суттєво відрізняються одна від одної, кожна має свої переваги та недоліки.
Класичні системи
Системи цієї групи базуються, в першу чергу, на морфологічних особливостях будови талому та колоній, а також на типах розмноження. Морфологічний підхід до систематики Cyanophyta був розроблений на початку ХХ ст. одразу кількома альгологами, серед яких особливо значну роль відіграли роботи Л. Гейтлера (Geitler) та О.О. Єленкіна. Класичні системи зручні у роботі, дозволяють порівняно легко визначати види, знаходити місце у системі новим, раніш не відомим таксонам. Головним недоліком класичних систем є те, що вони майже не відображають реальну філогенію відділу.
За класичними системами відділ найчастіше поділяли на три класи - Chroococcophyceae, Chamaesiphonophyceae та Hormogoniophyceae. Основними ознаками класів були план будови вегетативного тіла (одноклітинний чи багатоклітинний), спосіб розмноження (зокрема, здатність до утворення наноцитів, екзоцитів, гормогоніїв та гормоцитів), частково - спосіб життя (прикріплений чи вільний).
Морфолого-цитологічна система
Система була розроблена Й. Комареком та К. Анагностидісом (Komarek, Anagnostidis) у середині 80-х років ХХ ст. з метою узагальнення численних даних про цитологію окремих представників та ряду результатів порівняльних фізіолого-біохімічних досліджень Cyanophyta. Таксони рангу класу та окремих порядків добре співпали з молекулярно-філогенетичними деревами. Проте на рівні середніх таксонів - родин та родів, система лишилася вельми штучною. Крім того, використання ряду ультраструктурних ознак як діагностичних критеріїв родин та родів зробило її більш складною у практичній роботі порівняно з морфологічною системою. Незважаючи на це, саме морфолого-цитологічна система сьогодні стала провідною у практиці світових альгологічних досліджень.
Згідно з морфолого-цитологічною системою, відділ Cyanophyta включає лише один клас - Cyanophyceae, що поділяється на чотири порядки - Chroococcales, Oscillatoriales, Nostocales та Stigonematales (табл. 9.1).
Таблиця 9.1. Ознаки порядків класу Cyanophyceae
Порядок |
План будови |
Тип поділу клітин |
Гетероцисти та акінети |
Справжнє галуження |
Chroococcales |
одноклітинний |
повний |
відсутні |
відсутнє |
Oscillatoriales |
багатоклітинний |
неповний |
відсутні |
відсутнє |
Nostocales |
багатоклітинний |
неповний |
наявні |
відсутнє |
Stigonematales |
багатоклітинний |
неповний |
наявні |
наявне |
Chroococcales включає всі одноклітинні синьозелені водорості. Поділ клітин повний, основним типом є бінарний поділ, додатковими - рівний багаторазовий та нерівний поділи з утворенням наноцитів та екзоцитів. Хроококальні водорості можуть бути представлені поодинокими клітинами або утворювати різноманітні колонії.
Найбільш примітивною синьозеленою водорістю вважається Gloeobacter, що мешкає у гарячих джерелах. Клітини цієї водорості поодинокі, більш-менш циліндричні. Поділ відбувається лише у площині, перендикулярній до поздовжньої вісі клітини. Клітини глеобактеру не мають тилакоїдів та справжніх фікобілісом: хлорофіл та фікобілінові пігменти розташовуються безпосередньо на плазмалемі. Виявлено також деякі біохімічні особливості, що можна розглядати як ознаки примітивності: зокрема, водорость не синтезує деякі ліпіди (з групи діацилгліцеролів), що властиві всім іншим оксигенним фотосинтезуючим організмам, включаючи евкаріот. Припущення про примітивність глеобактеру повністю узгоджуються з молекулярно-біологічними даними.
Морфологічним "двійником" глеобактеру є рід Synechococcus. Проте клітини цієї водорості вже мають тилакоїди, типові фікобілісоми, синтезують діацилгліцероли. Представники роду зустрічаються у континентальних водоймах (включаючи термальні екотопи) та у морях. У прісноводної водорості Tubiella клітини подібні до глеобактеру та синехококу, проте знаходяться у трубчастих слизових колоніях (рис. 9.5).

Рис. 9.5. Хроококальні водорості. 1 - Synechococcus; 2 - Tubiella; 3 - послідовні стадії поділу клітин Synechocystis; 4 - Merismopedia; 5, 6 - Microcystis (5 - послідовні стадії поділу клітин, 6 - колонії); 7 - Gloeocapsa; 8, 9 - Chlorogloea (8 - колонія з кубічних агрегатів, 9 - глеокапсоподібний кубічний агрегат); 10 - фрагмент колонії Siphononema (за Кондратьєва, Коваленко, Приходькова, 1984; Kovacik, 1988).
Види роду Synechocystis мають більш-менш сферичні клітини, що зазвичай не утворюють колоній. Клітини діляться почергово у двох площинах, причому площина кожного наступного поділу проходить перпендикулярно до площини попереднього. Це можна добре спостерігати у тих випадках, коли водорость вирощується на агаризованому поживному середовищі. Аналогічні клітини та тип поділу властивий видам роду Merismopedia. Проте клітини мерисмопедії завжди утримуються разом колоніальним слизом; колонії мають вигляд прямокутних одношарових пластинок, у яких клітини розміщуються правильними взаємно перпендикулярними рядами. Обидва роди мешкають у планктоні прісних водойм та приморських солоних озер.
Представники роду Microcystis також мають сферичні клітини і утворюють слизові колонії. Але поділ клітин відбувається у трьох площинах, причому площина кожного наступного поділу не завжди проходить перпендикулярно до попереднього. Як наслідок, колонії набувають неправильної форми. У більшості видів цього роду клітини містять газові вакуолі. Microcystis мешкає у планктоні прісних та солонуватоводних водойм, деякі види (M. aeruginosa, M. wesenbergii) здатні до масового розвитку і є одними з найнебезпечніших збудників "цвітіння" води. Окремі види зустрічаються у морях, ґрунтах, на вологих аерофітних субстратах.
Клітини Gloeocapsa більш-менш сферичні, діляться у трьох взаємно перпендикулярних площинах, здатні до швидкого багаторазового поділу з утворенням наноцитів і утворюють характерні слизові колонії, що складаються з системи вкладених один в одного слизових міхурів. Види роду зустрічаються в біотопах всіх типів (морські та континентальні води, ґрунти), проте найчастіше оселяються на зрошуваних бризками скелях, стінах водоспадів, сирому камінні.
Масивні, звичайно макроскопічні слизові колонії, в яких клітини розміщуються нечіткими рядами, властиві родам Chlorogloea та Siphononema. В обох представників клітини діляться у трьох напрямках, здатні утворювати наноцити. Відміни полягають у тому, що клітини Chlorogloea звичайно позбавлені виразних індивідуальних слизових обгорток, через що колоніальний слиз здається безструктурним. Один з видів роду - Chlorogloea sarcinoides, масово розвивається у солоних озерах і бере участь в утворенні лікувального мулу - пелоїдів, інші види зустрічаються зрідка, переважно у перифітоні прісних водойм. У Siphononema індивідуальні клітинні обгортки чіткі, і колонії нагадують ниткоподібні комплекси з багатьох глеокапс. Водорость зрідка зустрічається у перифітоні чистих гірських струмків та річок.
Виразно ниткоподібні колонії утворюють Pascherinema (=Endonema), Hyella та Pleurocapsa. Хоча всі клітини здатні до поділу у кількох напрямках, проте звичайно поділ відбувається у площині, паралельній до площини попереднього поділу. Клітини у нитках утримуються разом тонкими та міцними слизовими піхвами. Досить часто утворюються наноцити. У Pascherinema колонії звичайно нерозгалужені, у Hyella слабо розгалужені. Колонії Pleurocapsa галузяться рясно, і місцями набувають паренхіматозного вигляду (рис. 9.6).
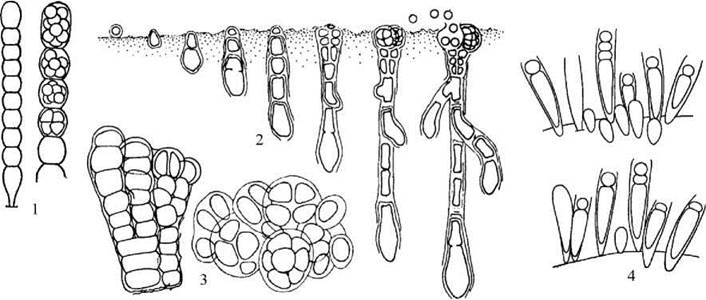
Рис. 9.6. Хроококальні водорості. 1 - Pascherinema: ниткоподібна колонія та утворення наноцитів; 2 - послідовні стадії розвитку та утворення наноцитів у свердлячої водорості Hyella; 3 - паренхіматозно-ниткоподібна колонія та утворення наноцитів у Pleurocapsa; 4 - Chamaesiphon: дорослі клітини, утворення екзоцитів та їх проростання (за Кондратьєва, Коваленко, Приходькова, 1984; Komarek, Anagnostidis, 1986).
Pascherinema зрідка зустрічається у прісних водоймах в обростаннях вищих водних рослин, а також на листках сфагнових мохів. Hyella належить до групи т.з. свердлячих водоростей: вона оселяється на вапнякових субстратах та вростає в них, спричинюючи їх руйнування. Pleurocapsa має широку екологічну амплітуду - у межах роду відомі прісноводні, морські, ґрунтові та аерофітні види.
Прикладом водорості, що здатна утворювати екзоцити є Chamaesiphon. Клітини хамесифону не утворюють колоній, оточені тонкою міцною слизовою піхвою, яка по завершенні поділу розривається на верхівці, звільнюючи екзоцит. Водорость мешкає у прісних водоймах, переважно - на нитчастих зелених водоростях.
Oscillatoriales об’єднує багатоклітинні гомоцитні водорості, що мають нерозгалужені трихоми. Поділ клітин є неповним і відбувається у площині, перпендикулярній поздовжній вісі трихому. Розмноження здійснюється гормогоніями або гормоцитами.
Прикладами примітивних осцилаторіальних є Pseudanabaena та Leptolyngbya. (рис. 9.7). Обидва роди мають тонкі трихоми (0.5-2 мкм завширшки), більш-менш діжкоподібні клітини, довжина яких приблизно вдвічі перевищує ширину. Тилакоїди розташовуються на периферії клітини кількома концентричними колами (рис. 9.8). Будь-яка клітина трихому здатна до поділу, причому перед кожним новим поділом дочірня клітина доростає до розміру материнської. Розмноження здійснюється рухливими гормогоніями, що утворюються шляхом фрагментації трихому. Види цих родів здатні до хроматичної адаптації.
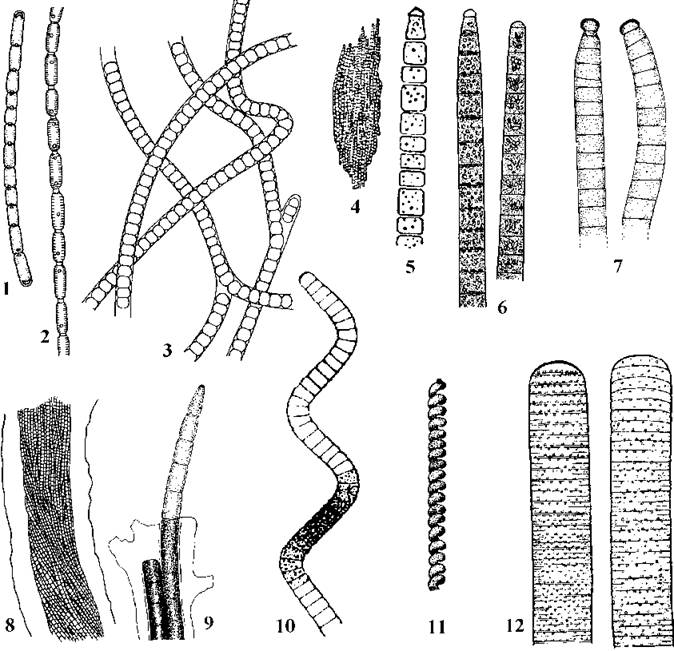
Рис. 9.7. Осцилаторіальні водорості. 1, 2 - Pseudanabaena (1 - P.galeata, 2 - P.catenata); 3 - Leptolyngbya boryana; 4, 5 - Trichodesmium erythraeum (4 - колонія, 5 - трихом); 6 - Planktothrix agardhii; 7 - Phormidiumautumnale; 8, 9 - Microcoleus vaginatus (8 - центральна частина нитки з багатьма трихомами, 9 - трихоми у верхівковій частині нитки); 10 - Arthrospira platensis; 11 - Spirulina major; 12 - Oscillatoria limosa (за Кондратьєва, 1969; Anagnostidis, Komarek, 1988; Komarek, Lund, 1990; Garbacki & al., 1999; ориг.).
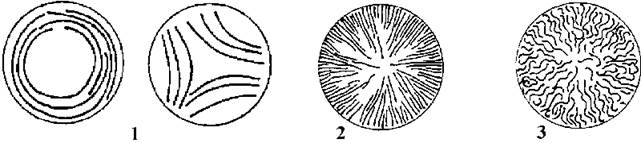
Рис. 9.8. Схема розміщення тилакоїдів у різних представників багатоклітинних синьозелених водоростей. 1 - Pseudanabaena, Leptolyngbya; 2 - Trichodesmium, Planktothrix, Phormidium, Microcoleus; 3 - Oscillatoria, Scytonema, Anabaena, Stigonema (за Anagnostidis, Komarek, 1988; 1990; Komarek, Anagnostidis, 1989).
Pseudanabaena має глибоко перетягнуті клітини, що містять одну-дві газові вакуолі. Трихоми порівняно короткі (до 50-80 клітин), позбавлені слизових піхв. Представники поширені у морях та прісних водоймах.
Leptolyngbya об’єднує тонконитчасті види, що за класичною системою входили переважно до родів Phormidium, Lyngbya, Plectonema. Лептолінгбії звичайно мають слизові піхви, досить часто (особливо у культурі) - несправжнє галуження і позбавлені газових вакуолей. Цікаво, що в деяких випадках у клітин спостерігається нерівний поділ, що нагадує поділ Chamaesiphon. Види роду широко розповсюджені переважно у бентосних та перифітонних угрупованнях прісних водойм, а також у ґрунтах. Кілька представників знайдено у морських біотопах.
Численну групу складають водорості, що мають трихоми середнього розміру (2-8 (15) мкм завширшки), циліндричні, неперетягнуті біля поперечних перегородок клітини, довжина яких у дорослому стані приблизно дорівнює ширині. Тилакоїди розташовуються на периферії клітини радіально. Будь-яка клітина трихому здатна до поділу, перед кожним новим поділом дочірня клітина доростає до розміру материнської. Після поділу між клітинами лишаються досить великі пори - близько 15-20 нм). Здатність до хроматичної адаптації у межах цієї групи не виявлена. Характерними представниками є роди Trichodesmium, Planktothrix, Phormidium, Microcoleus, Arthrospira.
Види роду Trichodesmium мешкають переважно у морському планктоні (зокрема, T. erythraeum спричинює "цвітіння" води у Червоному морі, через яке останнє отримало свою назву). Трихоми водоростей цього роду звичайно зібрані у колонії, в яких утримуються разом аморфним слизом, а клітини виповнені численними газовими вакуолями. Види роду Planktothrix за морфологією подібні до Trichodesmium, проте мешкають у планктоні прісних водойм і не утворюють слизових колоній. Ці водорості також викликають "цвітіння" води: наприклад, P. agardhii - в озерах та ставках рівнинних територій, P. rubescens - у гірських озерах (зокрема, в озерах Швейцірії "цвітіння", викликане цим видом зареєстровано ще у 1825 р.).
Phormidium - найбагатший за видами рід не тільки порядку, а й відділу в цілому (об’єднує більш 300 видів). На відміну від Trichodesmium та Planktothrix, клітини не мають газових вакуолей і мешкають у бентосі та планктоні морів та прісних континентальних водойм, а також у ґрунтах. Трихоми звичайно одягнені слизовими піхвами або позбавлені їх.
Для видів роду Microcoleus характерна наявність слизових піхв, в яких розміщується по кілька трихомів. За іншими ознаками Microcoleus нагадує попередній рід, і мешкає переважно у ґрунтах.
Характерною ознакою роду Arthrospira є наявність формідум- або триходесміум-подібних трихомів, які скручені у вигляді правильної широкої спіралі. Види роду зустрічаються у прісних водоймах з високим вмістом карбонатів (наприклад, A. platensis, A. maxima) або забруднених органічною речовиною (A. jenneri). Перші два види введені у промислову культуру і вирощуються у багатьох країнах світу як харчові водорості та фармакологічна сировина. Близьким до Arthrospira за морфолого-цитологічною системою вважається також рід Spirulina. Трихоми у видів цього роду тонкі, скручені у дуже щільні спіралі. Мешкають види Spirulina переважно у морях, приморських лиманах, рідше зустрічаються у прісних континентальних водоймах.
Прикладом іншої морфолого-цитологічної групи є рід Oscillatoria. Трихоми водорості звичайно широкі (6-60 мкм завширшки), інколи знаходяться у слизових піхвах, складаються з дуже коротких дископодібних клітин. Тилакоїди викривлені, виповнюють майже всю порожнину клітини. Для Oscillatoria та споріднених з нею родів властивий особливий тип поділу клітин та утворення гормогоніїв та гормоспор. Поділ клітин відбувається лише у певних ділянках трихому, що отримали назву меристемних зон. При цьому поділ кожної клітини відбувається дуже швидко, і нові поперечні клітинні перегородки починають закладатися ще до закінчення попереднього поділу. Пори у поперечних перегородках дуже дрібні у порівнянні з іншими осцилаторіальними (близько 5 нм у діаметрі). Гормогонії та гормоцити утворюються після відмирання окремих клітин трихому. Ці клітини називають некридіями. Види роду зустрічаються найчастіше у приморських лиманах та прісних континентальних водоймах.
Nostocales об’єднує багатоклітинні гетероцитні водорості з нерозгалуженими трихомами. Як і у Oscillatoriales, поділ клітин є неповним і відбувається у площині, перпендикулярній поздовжній вісі трихому, а розмноження здійснюється гормогоніями або гормоцитами.
Ностокальні водорості поширені у морях, континентальних водоймах, у ґрунтах, і завдяки здатності до фіксації атмосферного азоту в аеробних умовах відіграють надзвичайно важливу роль у планетарному кругообігу азоту. Деякі види ностокальних є небезпечними збудниками "цвітіння" води.
Примітивну лінію ностокальних представляє рід Scytonema. Ця водорость має довгі, однакові за шириною трихоми, оточені досить міцними слизовими піхвами. Поділ клітин неповний, дещо нагадує поділ у Oscillatoria. Між вегетативними клітинами трихому розташовується інтеркалярні гетероцисти. Нитки здатні до подвійного (т.з. сцитонемоїдного) галуження, що виникає, як правило, внаслідок відмирання однієї або кількох вегетативних клітин. При цьому трихоми продовжують рости, тиснуть один на одного, і кінець кінцем проривають материнську піхву, утворюючи дві бічні несправжні гілки. Види роду найчастіше зустрічаються у наземних біотопах, і особливо численні у субтропічних та тропічних регіонах та тундрових біогеоценозах (рис. 9.9).
Більш продвинутою в еволюційному відношенні є Gloeotrichia. Водорость утворює масивні слизові колонії, шо складаються з нерозгалужених асиметричних ниток. При основі трихому знаходиться базальна гетероциста, за нею (у зрілих індивідів) - видовжена акінета, далі - вегетативні клітини. Останні біля гетероцисти широкі та короткі, при наближенні до верхівки звужуються та витягуються у довжину, і на самій верхівці закінчуються довгим безбарвним волоском. Gloeotrichia мешкає переважно у прісних стоячих водоймах. На початку розвитку колонії прикріплені до підводних субстратів (переважно до стебел та листків вищих водних рослин), згодом відриваються від них і вільно плавають на поверхні води. У деяких країнах Азії місцеві жителі збирають колонії глеотрихії та вживають у їжу.
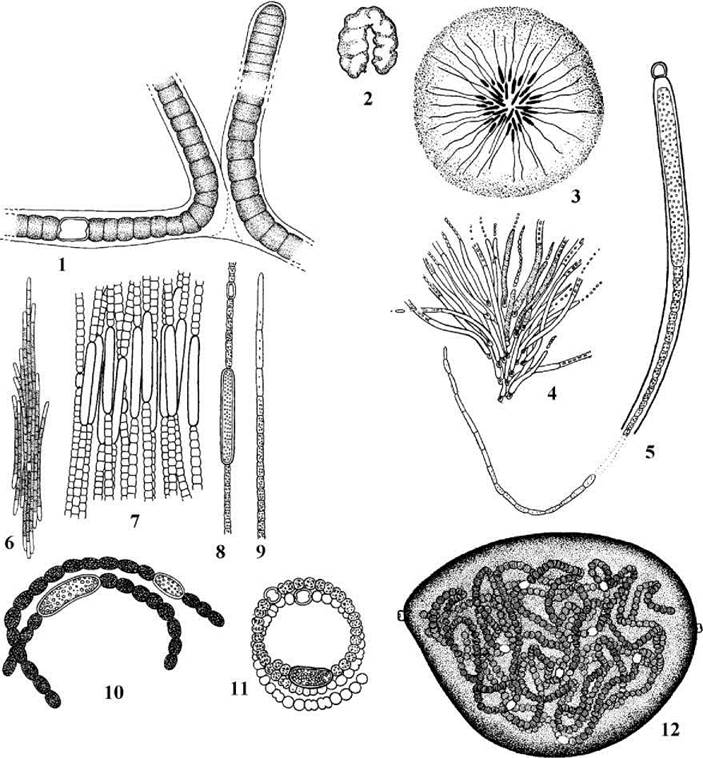
Рис. 9.9. Ностокальні водорості. 1 - Scytonema julianum; 2-5 - Gloeothrichia intermedia (2 - зовнішній вигляд колонії, 3 - розташування ниток у колоніальному слизу, 4 - фрагмент колнії з групою ниток, 5 - нитка з гетероцистою, акінетою, волоском); 6-9 - Aphanizomenon flos-aquae (6 - колонія, 7 - центральна частина колонії, 8 - центральна частина трихому з гетероцистою та акінетою, 9 - верхівка трихому з безбарвною клітиною); 10, 11 - Anabaena flos-aquae; 12 - Nostoc commune (за Кондратьєва, 1969; Garbacki & al., 1999).
Водорості, що мають симетричні нерозгалужені нитки вважаються вершиною еволюції порядку. Характерні приклади цієї гілки - роди Aphanizomenon, Anabaena, Nostoc.
Трихоми Aphanizomenon з'єднуються у шкуринки, що плавають у товщі води. Кожний трихом дещо звужується на верхівках і закінчується безбарвними витягнутими клітинами. Центральна частина трихому утворена вегетативними клітинами, між якими розташовуються поодинокі інтеркалярні гетероцисти та акінети. Клітини виповнені численними газовими вакуолями. Всі види роду є небезпечними збудниками токсичного "цвітіння" води. Так, у водосховищах Дніпровського каскаду приблизно 20% випадків "цвітіння" зумовлює A. flos-aquae. Окремі форми цього виду також спричинюють "цвітіння" води у Азовському морі (починаючи з середини 90-х років ХХ ст. це "цвітіння" спостерігається регулярно).
Трихоми Anabaena поодинокі, колоній не утворюють, досить часто згорнуті у щільну спіраль. У багатьох видів клітини містять газові вакуолі. Види роду мешкають переважно у планктоні прісних водойм. Деякі види Anabaena (зокрема, A. flos-aquae, A. spiroides, A. scheremetievi) належать до збудників токсичного "цвітіння" води.
До роду Nostoc належать водорості із симметричними трихомами, що розташовуються у масивних слизових колоніях. Клітини ностоку позбавлені газових вакуолей, біля поперечних перегородок звичайно дуже перетягнуті. Мешкають у прісних водоймах та ґрунтах. Види з крупними колоніями використовують в їжу.
Stigonematales об'єднує гетероцитні водорості з розгалуженими трихомами. Представники порядку, що морфологічно не відрізняються від сучасних, відомі з кембрію (570-500 млн. років). Клітини стигонематальних здатні до поділу у кількох площинах, внаслідок чого трихоми утворюють справжні гілки, а іноді - навіть паренхіматозні структури, що нагадують справжні тканини. Мешкають стигонематальні переважно у тропічних та субтропічних регіонах. На території України з цього порядку досить часто зустрічається лише рід Stigonema (рис. 9.10).
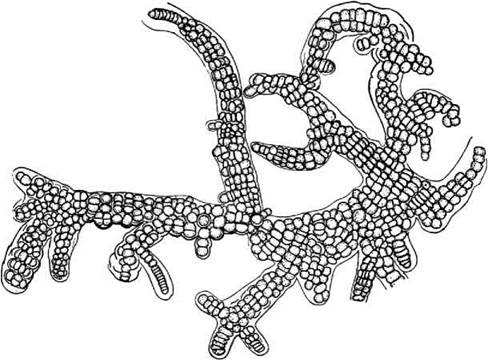
Рис. 9.10. Stigonema intermedia: зовнішній вигляд зрілої розгалуженої нитки (Кондратьєва, 1969).
Молекулярно-біологічна система
Перші спроби застосувати молекулярно-біологічні методи для побудови системи синьозелених водоростей, максимально наближеної до природньої, були здійснені у середині 80-х років ХХ ст. У 90-х років ці дослідження набули широкого масштабу, і внесли суттєві корективи у погляди на основні напрямки еволюції Cyanophyta.
Основою молекулярно-біологічних досліджень у систематиці став метод побудови молекулярно-філогенетичних дерев за результатами аналізу нуклеотидних послідовностей гену, що кодує малу субодиницю (SSU) рибосомальної РНК. Цей ген був обраний не випадково - він представлений у всіх живих організмів (за винятком вірусів), відповідає за здійснення універсального для клітин процесу - біосинтезу білку, містить як досить консервативні, так і вельми мінливі ділянки. Завдяки останньому SSU у всіх організмів, з одного боку, виконує однакову універсальну функцію, а з іншого - має певні відміни у нуклеотидних послідовностях мінливих ділянок, за якими можна розраховувати філогенетичну відстань між різними представниками. Крім SSU, молекулярно- філогенетичні реконструкції проводили і за іншими генами та амінокислотними послідовностями різних білків (загалом біля 20).
Головною перевагою молекулярно-біологічної системи Cyanophyta є те, що сьогодні вона найбільш наближена до природньої системи відділу і дозволяє встановити основні напрямки еволюції синьозелених водоростей. Головними недоліками цієї системи є те, що по-перше, за молекулярними ознаками досліджено порівняно небагато видів (на 2000 р. - біля 400, тобто лише 20% від кількості відомих), по-друге, при побудові молекулярно-філогенетичних дерев біологи не дотримуються Кодексу ботанічної номенклатури і оперують не таксонами, а кладами та групами. Через це молекулярні системи на перший погляд здаються досить хаотичними, і робота з ними потребує попереднього детального знайомства з конкретними родами, видами та навіть окремими штамами, причому за системами, що ґрунтуються не на генотипічних, а класичних фенотипічних ознаках.
Дещо спрощений варіант молекулярно-філогенетичного древа Cyanophyta наведений на рис. 9.11).
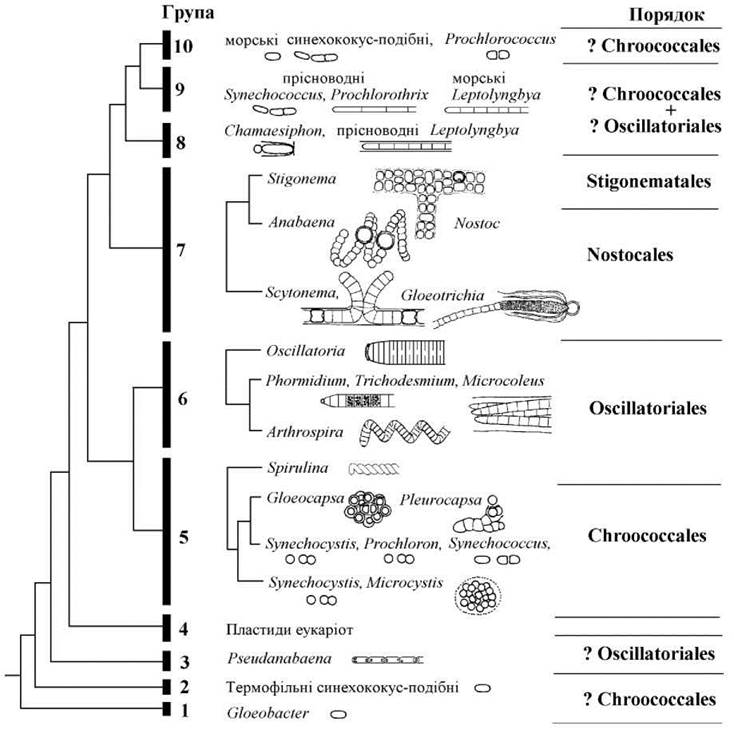
Рис. 9.11. Молекулярно-філогенетичне дерево Cyanophyta, побудоване за 16S РНК-кодуючим геном, та його узгодження з морфолого-цитологічною системою. Знак питання показує групи, належність яких до наведеного порядку є сумнівною (за основу взято дані Wilmotte, 1994; Turner, 1997).
За молекулярно-філогенетичним древом переважна більшість Cyanophyta увійшла в п’яту, шосту та сьому групи, які в цілому узгоджуються з порядками, виділеними за морфолого-цитологічною системою.
Аналіз молекулярно-філогенетичного дерева також показує кілька еволюційних тенденцій у межах відділу. По-перше, примітивні групи Cyanophyta (перша та друга) об’єднують одноклітинні термофільні види, більш продвинуті групи мають досить широку екологічну амплітуду. По-друге, багатоклітинність у синьозелених водоростей виникала неодноразово, зокрема у третій, п’ятій, восьмій та дев’ятій групах, у деяких випадках відбувався вторинний перехід від багатоклітинної будови до одноклітинної (десята група). По-третє, у різних групах Cyanophyta неодноразово з’являлися додаткові хлорофіли (зокрема, хлорофіл b) та хлорофіл-подібні пігменти (дивініл феопорфірин а5). Так, хлорофіл b наявний у водоростей родів Prochloron (група 5) та Prochlorothrix(група 9), хлорофіл-подібний пігмент - у Prochlorococcus (група 10). По-четверте, саме синьозелені водорості дали початок пластидам евкаріотичних організмів (група 4).
Прохлорофітові водорості та походження пластид
До середини 80-х років ХХ ст. вважали, що всі прокаріотичні оксигенні фотоавторофні організми мають лише один хлорофіл а та обов'язковий комплекс фікобілінових пігментів і належать до відділу Cyanophyta. У 1975 р. у субтропічних морських прибережних водах було знайдено одноклітинну прокаріотичну водорость з зеленим забарвленням, описану як новий для науки вид Synechocystis didemni. Водорость мешкала у симбіозі з асцидіями, розвиваючись у їх клоакальних порожнинах, покривах, на поверхні колоній. При біохімічних дослідженнях складу пігментів з'ясувалось, що клітини S. didemni мають два хлорофіли - а та b, і позбавлені фікобілінів. Продуктом асиміляції виявився полісахарид, подібний до справжнього крохмалю. Оскільки склад хлорофілів на той час вважався найсуттєвішою ознакою на рівні відділу, цю водорость було перенесено у новий рід Prochloron з єдиним видом P. didemni і виділено у новий самостійний відділ т.з. прокаріотичних зелених водоростей - Prochlorophyta (рис. 9.12).
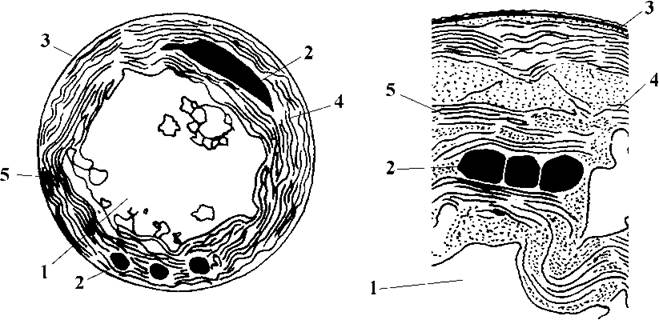
Рис. 9.12. Будова Prochloron за даними електронної мікроскопії. 1 - вакуолеподібна структура, 2 - поліедральне тіло, 3 - двошарова муреїнвмісна оболонка, 4 - поодинокий тилакоїд, 5 - група щільно притиснутих тилакоїдів, що нагадує ламелу. Ліворуч зображено клітину в цілому, праворуч її частину на великому збільшенні (схематизовано за Lewin, 19987, 1994).
Приблизно через десять років у планктоні озер у Нідерландах було знайдено багатоклітинну вільноіснуючу прокаріотичну водорость, що також містила хлорофіли а та b і не мала фікобілінів. Цей організм, що отримав назву Prochlorothrix hollandica, також було віднесено до Prochlorophyta.
Ще через два роки у пробах морського планктону відкрили нову групу прохлорофітових, представлену дрібними (0.6-0.8 мкм у діаметрі) прокаріотичними водоростями, які, хоча і не мали хлорофілу b, проте разом із хлорофілом а містили хлорофіл а-подібний пігмент дивініл феопорфірин а5. Фікобілінові пігменти також були відсутні. Види, що мали ці ознаки увійшли до роду Prochlorococcus.
Прохлорофітові водорості у 80-90-х стали об'єктами інтенсивних і численних біохімічних та електронно-мікроскопічних досліджень, оскільки припускалося, що саме серед Prochlorophytaзнаходилися ті організми, що вступивши у внутрішньоклітинний симбіоз з примітивними гетеротрофними евкаріотами, дали початок справжнім хлоропластам3.
У 1990-96 рр. гіпотезу прохлорофітного походження пластид евкаріотичних рослин перевіряли одразу кілька дослідницьких груп, що займалися питаннями молекулярної філогенії. Основні роботи проводилися методом побудови молекулярно-філогенетичних дерев за результатами аналізу генів, що кодують 16S субодиницю рибосомальної РНК (див. рис. 11.11), один з ферментів реакційного центру фотосистеми ІІ (psbA) та білки світозбираючої системи (LHCs).
В результаті цих робіт кількома групами незалежно було зроблено два головних висновки. По-перше, водорості, об'єднані у відділ Prochlorophyta, не є філогенетично спорідненими, і входять до різних груп Cyanophyta (п’ятої, дев’ятої та десятої). Оскільки порушується принцип монофілії відділу, відділ Prochlorophyta має бути спростованим.
По-друге, жодна з т.з. "прохлорофітових" водоростей не належить до пращурної групи пластид евкаріот. Пластиди походять від синьозелених водоростей середнього ступеню еволюційної продвинутості, що складали самостійну молекулярно-філогенетичну лінію в межах Cyanophyta. Сучасні види синьозелених водоростей з цієї групи невідомі.
Дослідження "прохлорофітових" водоростей також спростували положення про те, що в ході еволюції різні типи хлорофілів не могли виникати багаторазово.
Поширення та значення у природі та житті людини
Розповсюджені Cyanоphyta надзвичайно широко: в морях, прісних та, гіпергалійних водоймах, у ґрунтах, на снігу та льоду, в гарячих джерелах, в аерофітних умовах та ін. За стійкістю до дії екстремальних факторів Cyanоphyta посідають перші місця на планеті. Так, у модельних експериментах синьозелені водорості зберігали життєздатність у діапазоні температур від -195 до +130 °С, тиску від 0.05 до 300 атм., витримували опромінювання радіоактивним 60Co у 160 тис. р на годину (Microcoleus vaginatus - до 1280 тис. р на годину). Надзвичайна витривалість синьозелених водоростей навіть привернула увагу екзобіологів: зокрема, в США з початку 60-х років ряд лабораторій (наприклад, "Jet Propulsion Laboratory" в Каліфорнії) розробляють методики та ведуть селекцію штамів синьозелених водоростей, придатних розмножуватись у позаземних умовах, зокрема, на Марсі.
В наш час синьозелені водорості відіграють планетарну роль в азотному балансі. Це обумовлено здатністю представників Cyanophyta засвоювати азот безпосередньо з атмосфери. З діяльністю синьозелених водоростей-азотфіксаторів пов’язані явища відновлення родючості ґрунтів "під паром", збереження родючості цілинних ґрунтів, продуктивність біогідроценозів Світового океану.
Два види роду Arthrospira (A. platensis та A. maxima) є цінними біотехнологічними об’єктами. Вони введені у промислову культуру і їх біомаса використовується для виробництва дієтичних харчових продуктів, вітамінних домішок, фармакологічних препаратів (в першу чергу - радіопротекторів, стимуляторів обміну речовин, гормональних препаратів), харчових барвників тощо.
Синьозелені водорості використовують також для контролю якості води, в геологічній практиці при датуванні віку докембрійських осадових порід.
Серед Cyanоphyta є надзвичайно шкідливі види - збудники "цвітіння" води. Наприклад, у водосховищах Дніпровського каскаду влітку масово розмножуються Microcystis aeruginosa, Anabaenaflos-aquae, Aphanizomenon flos-aquae. При цьому різко погіршується кисневий режим, у воду потрапляють продукти розкладу відмерлих клітин, живі особини виділяють токсичні речовини. Факторами, що сприяють розвитку "цвітіння" є висока температура води (23-32 °С), висока концентрація біогенних елементів (азоту, фосфору, калію), відсутність перемішування водних мас.
Токсини синьозелених водоростей
За механізмом дії токсини синьозелених водоростей відносять до трьох груп: гепатотоксини, нейротоксини та дерматотоксини. Перші два типи розрізняють за клінічною картиною загибелі мишей при внутрішньому введенні досліджуваного токсину у летальних дозах при стандартних тестах. Гепатотоксини (т.з. фактори швидкої смерті) спричинюють прогресуючий цироз печінки і викликають загибель дослідної тварини протягом 45 хв. - кількох годин. Нейротоксини (фактори дуже швидкої смерті) у гострій дозі спричинюють смерть протягом 2-30 хв. внаслідок порушення дихальної функції. Дерматотоксини викликають гості дерматити при поверхневому контакті.
Сьогодні у Cyanophyta виявлено три основні гепатотоксини: мікроцистін, нодулярин (обидва є низькомолекулярними токсинами пептидної природи) та алкалоїд циліндроспермопсин. Мікроцистін виробляють Microcystis aeruginosa, Anabaena flos-aquae, Planktothrix agardhii. Нодулярин та циліндроспермопсин виявлені у кількох видів ностокальних водоростей.
З нейротоксинів для людини найбільшу небезпеку становлять алкалоїди групи анатоксинів та сакситоксинів. Серед збудників "цвітіння" води головними продуцентами анатоксинів є планктонні види роду Anabaena (в першу чергу - A. flos-aquae), та деякі Phormidium (зокрема, Ph. formosum). Сакситоксини синьозелених водоростей відомі під назвою токсин молюскового паралічу. Цей токсин нешкідливий для холоднокровних, зокрема, молюсків, проте здатний накопичуватися у їх тканинах. Споживання таких молюсків теплокровними призводить до отруєння. У прісних та солонуватих водоймах головним продуцентом сакситоксину серед Cyanophyta є Aphanizomenon flos-aquae, а у морях - види роду Trichodesmium.
Дерматотоксини (аплізіотоксин, лінгбіотоксин А) знайдені у деяких морських осцилаторіальних.
1 Застаріла назва наноцитів - ендоспори.
2 Застаріла назва екзоцитів - екзоспори.
3 Про історію дослідження Prochlorophyta, дискусії щодо їх місця у системі та ролі у походженні пластид у цей період див.: Н.В. Кондратьева. Прокариотические зеленые водоросли - Prochlorophyta (обзор литературных данных). Альгология, 1991, т. 1, № 3, с. 87-101.