Альгологія - Костіков І.Ю. - 2009-2013
Розділ 8. Життєві цикли
Зміни життєвих форм, які відбуваються з індивідом на протязі його онтогенезу складають життєвий цикл. Життєвий цикл охоплює всі стадії розвитку індивіду між однаковими життєвими формами (наприклад, від зиготи до зиготи, або від гамети до гамети, від спорофіту до спорофіту).
Типи життєвих циклів
Звичайно розрізняють два основних типи життєвих циклів: життєві цикли водоростей, позбавлених статевого процесу, та життєві цикли водоростей, які мають статевий процес. Водорості, які позбавлені статевого процесу, називають агамними, а їх життєві цикли - цикломорфозом. У водоростей, що мають статевий процес - т.з. еугамних - звичайно розрізняють три основні типи життєвого циклу - гаплофазний, диплофазний та гаплодиплофазний.
Цикломорфоз. Прикладом цикломорфозу може бути життєвий цикл Chlorella. У цієї водорості представлено лише дві життєві форми: вегетативні клітини та апланоспори. Нерідко цикломорфози бувають досить складними. Наприклад, у більшості видів Chlamydomonas монадні вегетатитвні клітини утворюють зооспори, які проростають у нові монадні вегетативні клітини, при несприятливих умовах ці клітини перетворюються на акінети або переходять у нерухливий стан, досить часто зберігаючи при цьому здатність до розмноження за допомогою нерухомих гемізооспор; при покращенні умов існування нерухомі клітини здатні відновити джгутики і знов повернутися у монадний стан (рис. 8.1).
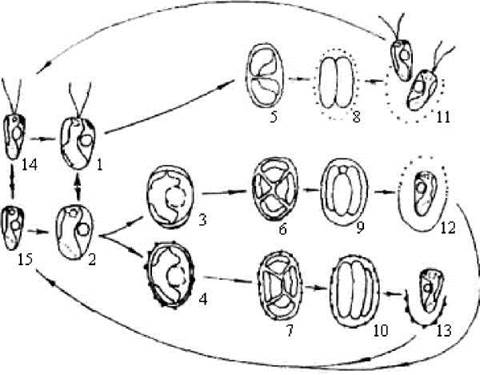
Рис. 8.1. Схема цикломорфозу агамної хламідомонади Chlamydomonas callunae: 1-3 - вегетативні клітини (1 - монадна, 2 - гемімонадна, 3 - кокоїдна); 4 - спочиваюча клітина - акінета; 5-7 - перетворення вегетативних клітин та акінети на спорангії; 8-10 - спорангії; 11 - вихід зооспор із спорангію, що утворився з монадної клітини; 12, 13 - звільнення гемізооспор зі спорангіїв, що утворилися з кокоїдної клітини або акінети; 14 - зооспора; 15 - гемізооспора.
Життєві цикли еугамних водоростей. Все ж таки коли мова йде про життєві цикли, звичайно мають на увазі цикли розвитку видів, що мають статевий процес. У життєвих циклах водоростей із статевим процесом обов'язково присутні принаймні три стадії: вегетативна стадія, гамети (або вегетативні клітини, що виконують їх функцію), зигота. Назву життєвим циклам дають, в першу чергу, за наборами хромосом (т.з. плоїдністю) вегетативних стадій резвитку. Якщо вегетативна стадія має гаплоїдний набір хромосом, життєвий цикл називають гаплофазним, диплоїдний набір - диплофазним. Якщо ж в життєвому циклі чергуються вегетативні стадії як з гаплоїдним, так і з диплоїдним набором хромосом, то життєвий цикл розглядається як гаплодиплофазний. Послідовність змін плоїдності протягом життєвого циклу називають зміною ядерних фаз.
Під вегетативними стадіями (поколіннями) розуміють ті стадії онтогенезу, на яких водорості активно вегетують та утворюють репродуктивні клітини. Під репродуктивними клітинами розуміють спеціалізовані клітини як нестатевого, так і статевого розмноження - спори та гамети. Залежно від типу розмноження (нестатеве або статеве) серед вегетативних стадій розрізняють спорофіт - покоління, що утворює спори, гаметофіт - покоління, що утворює гамети, та гаметоспорофіт - покоління, яке здатне утворювати як гамети, так і спори.
У життєвому циклі може бути присутнє тільки одне покоління (наприклад, тільки гаметофіт або тільки гаметоспорофіт), або різні покоління нерегулярно чи регулярно змінюються одне одним (наприклад, спорофіт змінюється гаметофітом, а гаметофіт, в свою чергу - спорофітом). Залежно від цього розрізняють життєві цикли без зміни поколінь, з неправильним чергуванням поколінь та з правильним чергуванням поколінь. Якщо при чергуванні поколінь різні покоління морфологічно подібні (наприклад, гаметофіт зовні не відрізняється від спорофіту), то таке чергування поколінь називають ізоморфним, у протилежному випадку (наприклад, коли спорофіт макроскопічний, а гаметофіт мікроскопічний) чергування поколінь називають гетероморфним.
Плоїдність поколінь, і, відповідно, тип життєвого циклу за чергуванням ядерних фаз, визначається тим, на який стадії життєвого циклу відбувається мейотичний (редукційний) поділ. При мейозі кількість хромосом зменшується вдвічі. Таким чином, всі стадії розвитку протягом відрізку життєвого циклу від мейозу до статевого процесу будуть мати гаплоїдний набір хромосом, а всі стадії після статевого процесу до мейозу - диплоїдний набір.
Типи мейозу та зміна ядерних фаз
Мейоз (редукційний поділ) може відбуватися на одній з трьох стадій життєвого циклу: 1) на стадії проростання зиготи - т.з. зиготична редукція, 2) на стадії утворення гамет (гаметична редукція), 3) при утворенні спор (спорична редукція) (рис. 8.2). Крім того, відомі випадки, коли мейоз відбувається у вегетативних (соматичних) клітинах (соматична редукція).
При зиготичній редукції вегетативне покоління завжди буде гаплоїдним, відповідно, життєвий цикл буде гаплофазним (наприклад, у Chlamydomonas moewusii). Звичайно перед мейозом зигота проходить період спокою, протягом якого відбувається підготовка до редукційного поділу (рис. 8.3: 1,2).
При гаметичній редукції вегетативне покоління завжди буде диплоїдним, відповідно, життєвий цикл - диплофазним (наприклад, у видів роду Fucus). В цих випадках зигота проростає, як правило, без періоду спокою. Таким чином, наявність періоду спокою дозволяє встановити тип мейозу, і тим самим визначити життєвий цикл. Виключення з цього правилу зустрічаються лише у динофітових водоростей (рис. 8.3: 3,4).
При споричній редукції диплоїдна зигота проростає без періоду спокою у диплоїдний спорофіт. На спорофіті утворюються спорангії, в яких відбувається мейоз і далі формуються гаплоїдні спори. Спори проростають у гаплоїдний гаметофіт, що продукує гамети. Після копуляції гамет утворюється диплоїдна зигота і цикл замикається. Оскільки при споричній редукції спорфіт є диплоїдним, а гаметофіт - гаплоїдним, життєвий цикл буде диплогаплофазним (рис. 8.3: 5).
Таким чином, тип редукції визначає тип життєвого циклу за ядерними фазами вегетативних поколінь: гаплофазний, диплофазний, диплогаплофазний.
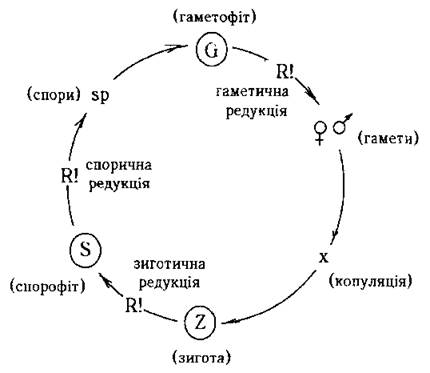
Рис. 8.2. Можливі типи мейозу (редукційного поділу) та умовні позначення основних стадій життєвого циклу.
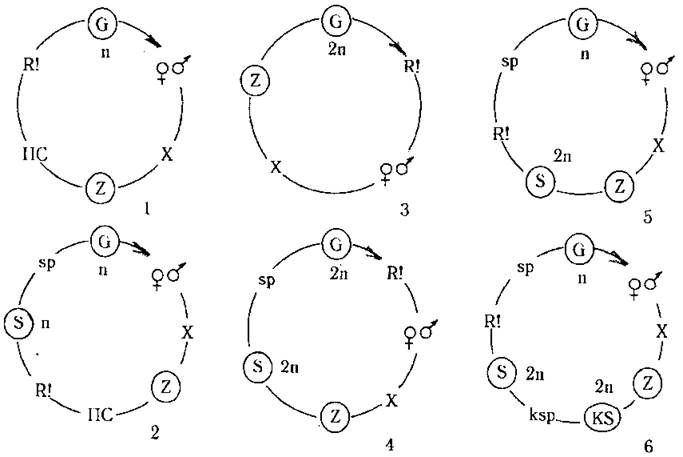
Рис. 8.3. Типи життєвих циклів у водоростей за зміною ядерних фаз та чергуванням поколінь: 1, 2 - гаплофазний цикл з зиготичною редукцією (1 - без чергування поколінь, 2 - з чергуванням поколінь); 3, 4 - диплофазний цикл з гаметичною редукцією (3 - без чергування поколінь, 4 - з чергуванням поколінь); 5, 6 - гаплодиплофазний цикл зі споричною редукцією (5 - з чергуванням двох поколінь - спорофіту та гаметфіту, 6 - з чергуванням трьох поколінь - гаметофіту, спорофіту та карпоспорофіту). Позначення: n - гаплоїдний набір хромосом, 2n - диплоїдний набір хромосом, KS - карпоспорофіт, ksp - карпоспори, ПС - період спокою. Інші позначення подані згідно до рис. 8.2.
Чергування поколінь
Біологічні передумови виникнення чергування поколінь. Для відтворення себе у майбутніх поколіннях особини будь-якого виду здійснюють кілька функцій: вони мають рости, займати нові території, підтримувати певний рівень генетичної різноманітності. Відповідно до цього розрізняють функцію основну - ростову, розселювальну та рекомбінантну.
Основна функція у водоростей реалізується переважно за рахунок фотосинтезу, а у безбарвних представників - за рахунок гетеротрофного живлення. Функція розселення здійснюється різними шляхами: у монадних та амебоїдних форм - через активний рух вегетативних клітин, у нерухомих представників - шляхом пасивного переносу вегетативних клітин, багатоклітинних таломів або їх фрагментів (течіями, вітром, тваринами тощо). Проте у переважної більшості мікроскопічних представників і значної частини макроскопічних форм функцію розселення виконують спеціалізовані клітини нестатевого розмноження - спори. Здійснення рекомбінантної функції досягається, в першу чергу, завдяки статевому процесу.
Гаметоспорофіт та життєві цикли без чергування поколінь. У водоростей часто зустрічаються варіанти, коли одне й те ж саме покоління виконує всі три функції: і росте, і утворює як спори, так і гамети. Таке "універсальне» покоління називають гаметоспорофітом (рис. 8.4: 1).
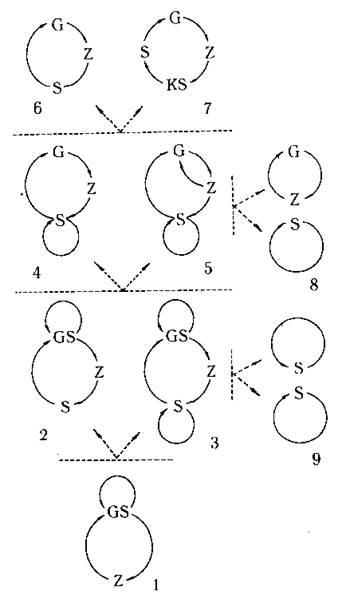
Рис. 8.4. Ускладнення життєвих циклів за чергуванням поколінь. 1 - цикл без чергування поколінь зі стадією гаметоспорофіту; 2, 3 - цикли з чергуванням гаметоспорофіту та спорофіту; 4, 5 - цикли з неправильним чергуванням гаметофіту та спорофіту (4 - моногенетичний, 5 - дигенетичний); 6 - цикл з правильним чергуванням гаметофіту та спорофіту; 7 - цикл з правильним чергуванням гаметофіту, карпоспорофіту та спорофіту; 8, 9 - цикли без чергування поколінь, що виникають внаслідок розімкнення циклів з чергуванням поколінь. GS - гаметоспорофіт, G - гаметофіт, S - спорофіт, KS - карпоспорофіт, Z - зигота.
Наприклад, саме такий варіант можна зустріти у одноклітинної зеленої водорості Chlorococcum: у неї зооспори водночас здатні виконувати і функцію гамет. Проте потенційна можливість здійснювати однією й тією ж самою клітиною водночас як розсілювальної, так і рекомбінантної функцій, призводять до певного конфлікту: з одного боку, спори потребують якомога більшого запасу поживних речовин, тому їх кількість не може бути великою; гамети ж, навпаки, в першу чергу мають бути численним, і через це не можуть містити достатньої кількості резервних асимілятів. Усереднення розмірів і кількості "універсальних" репродуктивних клітин призводить до зменшення ефективності виконання як розсілювальної, так і рекомбінантної функції.
Через це у більшості випадків гаметоспорофіт утворює два типи репродуктивних клітин, відмінних як функціонально, так і морфологічно: та числені гамети і великі, відносно нечисленні спори. Наприклад, у видів роду Chlamydomonas, які мають ізогамний статевий процес, одноклітинний монадний гаметоспорофіт може утворювати або 2-4 великі зооспори, або 8-16 дрібніших ізогамет.
Спорофіт та виникнення чергування поколінь. Подальша спеціалізація, яка підвищує ефективність виконання розсілювальної та рекомбінантної функцій, призводить до появи чергування поколінь: крім покоління, яке продовжує здійснювати рекомбінантну та розсілювальну функцію - гаметоспорофіту, з'являється нове покоління - спорофіт, яке реалізує лише наслідки вдалого акту запліднення (рис. 8.4: 2;3).
Гаметоспорофіт на утворення гамет витрачає значний відсоток запасів речовини та енергії. Проте як результат утворюється лише невелика кількість зигот, оскільки переважна більшість гамет не знаходять свого статевого партнера і гине. Наприклад, у зеленої водорості Volvox при приблизно рівній кількості антеридіїв та оогоніїв на одну яйцеклітину припадає 64 сперматозоїди. Зрозуміло, що 63 з них не приймуть участь у заплідненні і загинуть. У роду Ulva на один реалізований статевий акт припадає близько кількох тисяч гамет, що не прокопулювали; у роду Charaкількість сперматозоїдів, які формуються для запліднення однієї яйцеклітини, складає біля 40 тисяч. Таким чином, кожна зигота має надзвичайно високу "собівартість". Тому вигідною стратегією є не пряме проростання зиготи у гаметоспорофіт, а в спорофіт - особливе "заощадливе" покоління, яке і не витрачає обмежений ресурс на приречені гамети, а лише тиражує наслідки вдалого статевого процесу.
У найпростіших випадках спорофіт утворює спори, які надалі розвиваються в гаметоспорофіт (наприклад, у роду Ulothrix). Такі спорофіти зазвичай є одноклітинними (рис. 8.4: 2).
У складніших випадках спорофіт утворює спори, які надалі знов проростають у спорофіт - тобто відбувається багаторазове відтворення наслідків одного вдалого акту запліднення. І лише за певних умов (наприклад, стрибкоподібної зміни солоності, температури, концентрації певних речовин в оточуючому середовищі), спорофіт утворює спори, які проростають у гаметоспорофіт (рис. 8.4: 3).
Гаметофіт. Цикли з чергуванням спорофіту та гаметофіту. Наступне ускладнення життєвого циклу пов'язують з редукцією у гаметоспорофіту здатності до утворення спеціалізованих клітин нестатевого розмноження. Це призвело до появи покоління, спеціалізованого лише на забезпеченні рекомбінантної функції - гаметофіту. Цикли з чергуванням спорофіту та гаметофіту у водоростей можуть бути представлені варіантами з неправильним (неусталеним) або з правильним чергуванням поколінь.
При неправильному чергуванні поколінь спорофіт і гаметофіт змінюють один одного нерегулярно. Наприклад, спорофіт у бурої водорості Ectocarpus у багатогніздних спорангіях утворює зооспори, з яких знов виростають спорофіти, а в одногніздних - зооспори, які проростають у гаметофіт. Таким чином, спорофіт не завжди буде давати початок гаметофіту, що і визначає неусталеність чергування поколінь (рис. 8.4: 4).
Неправильна зміна поколінь може бути обумовлена також здатністю гамет проростати партеногенетично у нові гаметофіти. Таке явище досить часто зустрічається у бурих та сифональних зелених водоростей.
Крім того, неусталена зміна поколінь може бути наслідком поліваріантності шляхів розвитку зиготи. Наприклад, у морської водорості Bryopsis зигота може прорости або у спорофіт, або у гаметофіт (рис. 8.4:5). Цикл розвитку, що спостерігають у цьому випадку, отримав назву дигенетичного циклу. Фактори, що визначають характер проростання зиготи, лишаються незрозумілими.
Найбільш досконалими вважають цикли, при яких спостерігається повна спеціалізація поколінь за функціями і при яких спорофіти завжди дають початок тільки гаметофітам, і навпаки - тобто цикли з правильним чергуванням поколінь. Цікаво, що саме такі цикли притаманні всім вищим рослинам та тим водоростям, які сьогодні вважають еволюційно найбільш продвинутими (рис. 8.4: 6).
Життєві цикли з чергуванням трьох поколінь. Особливий тип життєвого циклу з правильним чергуванням поколінь представлений у червоних водоростей, де закономірно змінюють одне одного не два, а три вегетативних покоління - гаметофіт, карпоспорофіт та спорофіт (рис. 8.3: 6; 8.4: 7). Детально цей тип циклу розглядається у розділі, присвяченому червоним водоростям.
Розімкнення циклів та стрибкоподібне видоутворення
Сьогодні вже вважається доведеним, що трансформації життєвих циклів з неправильним чергуванням поколінь принаймні у деяких випадках призводили до стрибкоподібного видоутворення. Такі трансформації найчастіше відбувалися або внаслідок розімкнення життєвого циклу через вторинну втрату здатності до статевого відтворення (рис. 8.4: 8), або через втрату здатності зиготи проростати у спорофіт при дигенетичному шляху розвитку (рис. 8.4: 9).
Порушення циклів через втрату здатності до статевого розмноження. Класичний приклад подібного явища можна спостерігати у бурих водоростей з роду Cutleria. Так, у Середземному морі досить поширеним видом є Cutleria adspersa. Вона має диплогаплофазний цикл розвитку зі споричною редукцією та неправильним чергуванням спорофіту та гаметофіту. Гаметофіти однорічні, звичайно мають вигляд кущиків до 10-15 см заввишки. Спорофіти переважно багаторічні, мають вигляд розпростертої на субстраті товстої плівки, і раніш були описані як самостійний рід "Algaozonia".
Cutleria adspersa зустрічається також і в Чорному морі, проте лише у вигляді гаметофітів, і відтворює себе внаслідок партеногентичного проростання гамет. При цьому статевий процес не відбувається. В той же час у західній частині Середземного моря мешкають лише спорофіти Cutleria. Експериментальне було встановлено, що відтворюють вони себе за допомогою зооспор, при цьому мейоз перед утворенням зооспор не відбувається.
Таким чином, у межах Середземноморського басейну Cutleria adspersa існує у вигляді трьох популяцій, зв'язок між якими втрачений: чорноморська популяція, представлена лише гаметофітами (форма "Cutleria"), західно-середземноморська популяція, представлена лише спорофітами (форма "Algaozonia"), та східно-середземноморська популяція, представлена переважно спорофітами ("Algaozonia"), яка інколи ще дає стадії гаметофіту ("Cutleria"). Цей факт інтерпретується як приклад видоутворення, при якому стрибкоподібно виникають два нові, зовсім несхожі ані за морфологією, ані за біологією види, які, проте, є білизькоспорідненими.
Стрибкоподібне видоутворення внаслідок розмикання дигенетичних життєвих циклів можна бачити на прикладі Bryopsis. Для роду характерний диплофазний життєвий цикл з гаметичною редукцією. Причому в межах роду відомі види, у яких є чергування спорофіту та гаметофіту і дигенетичний тип проростання зиготи, так і види, представлені виключно гаметофітами.
Так, у Bryopsis halimeniae гаметофіт має вигляд системи пірчасте розгалужених "пагонів", а спорофіт - системи майже нерозгалужених ниток. Спорофіт раніш був описаний як самостійний вид роду Derbesia. Зигота, що утворюється після копуляції гамет, може проростати або у спорофіт ("Derbesia"), або у гаметофіт ("Bryopsis"). Спорофіт дає зооспори, які також можуть або відтворювати спорофітне покоління, або проростати у гаметофіт.
В той же час існують види роду Bryopsis, які не мають стадії спорофіту, і відтворюють себе лише статевим шляхом - тобто зигота завжди проростає лише в гаметофіт. Відомі також кілька видів роду Derbesia, які не мають статевого розмноження, і відтворюють себе тільки нестатевим шляхом - за допомогою зооспор. Вважають, що такі різні за розмноженням та морфологією представники як Bryopsis з моногенетичними циклам та справжні Derbesia, є близькоспорідненими, і виникли внаслідок втрати здатності зиготи розвиватися у спорофіт, тобто внаслідок розімкнення дигенетичного циклу з чергуванням поколінь.
Сьогодні трансформації життєвих циклів дозволяють знайти пояснення випадкам, коли при молекулярно-філогенетичних реконструкціях близькоспорідненими виявляються види, роди і навіть родини, що принципово різняться за морфологією та розмноженням, і за класичними системами розглядались як таксони неспоріднені або споріднені лише віддалено.
Ізоморфна та гетероморфна зміна форм розвитку
При наявності чергування поколінь гаметфіт (або гаметоспорофіт) та спорофіт можуть бути морфологічно подібними - ізоморфними, або суттєво відмінними - гетероморфними. Прикладом виду з ізоморфними поколіннями може бути морська зелена водорість Ulva: і спорофіт, і гаметофіт у неї мають вигляд складчастої по краям платівки і морфологічно не відрізняються (рис. 8.5: 1).

Рис. 8.5. Вегетативні стадії Ulva та Laminaria. 1 - Ulva (спорофіт та гаметофіт морфологічно не відрізняються один від одного і обидва мають вигляд макроскопічної зеленої платівки); 2-4 - Laminaria (2 - макроскопічний спорофіт, 3 - мікроскопічний жіночій гаметофіт, 4 - мікроскопічний чоловічій гаметофіт).
Класичним прикладом виду з гетероморфними поколіннями є Laminaria. Спорофіт ламінарії макроскопічний, має вигляд платівки з стеблоподібною нижньою частиною та ризоїдами; гаметофіт - мікроскопічний, має вигляд коротенької розгалуженої нитки (рис. 8.5: 2-4).
Єдиної точки зору стосовно того, яка зміна форм розвитку є первинною - ізоморфна чи гетероморфна, сьогодні немає. Проте існує одностайна думка, що саме гетероморфна зміна поколінь, "відпрацьована" у водоростевому світі, виявилась найбільш еволюційно вдалою, і саме з нею у значній мірі пов'язують макроеволюцію у світі вищих рослин.
Статевий диморфізм
Статевий диморфізм у водоростей, як і у інших груп еукаріот, полягає у морфологічних відмінах особин різних статевих знаків (наприклад, чоловічих та жіночих). Статева диференціація у водоростей - явище багатопланове, і простежується як на рівні гамет (або вегетативних клітин, що виконують функції гамет при соматогамних статевих процесах - див. розділ **), так і на рівні вегетативних поколінь, причому не тільки тих, що безпосередньо продукують гамети, а навіть на рівні спорофітів.
Види, у яких одна й та ж сама особина здатна продукувати гамети обох статевих знаків, називають однодомними або двостатевими. Наприклад, у водяної сіточки (Hydrodictyon) одна й та ж сама клітина продукує як "чоловічі", так і "жіночі" гамети - тобто є двостатевою (однодомною). Інший приклад однодомної рослини - водяна повсть (Vaucheria): у вошерії як сперматозоїди, так і яйцеклітини утворює одна й та ж сама рослина.
Види, у яких чоловічі та жіночі гамети утворюються на різних особинах, називають роздільностатевими (дводомними). Прикладом може бути Laminaria - у видів цього роду чоловічі гамети - сперматозоїди - утворюються на ниткоподібному чоловічому гаметофіті, який складається з дрібних клітин, а жіночі гамети - яйцеклітини - на жіночому гаметофіті, який складається з досить великих клітин (рис. 8.5: 3, 4).
Коли чоловіча та жіноча рослини помітно відрізняються за зовнішнім виглядом, то таке явище називають статевим диморфізмом. Наприклад, статевий диморфізм певним чином виражений у ламінарії, адже клітини жіночого та чоловічого гаметофіту різняться за розмірами.
У водоростей відомі випадки, коли жіночі та чоловічі особини абсолютно несхожі між собою, і тому статевий диморфізм проявляється надзвичайно яскраво. Зокрема, таке явище можна спостерігати у значної частини едогонієвих водоростей - серед видів, які належать до так званих нанандрієвих (від "нанос" -найдрібніший, та "андрос" - чоловік) (рис. 8.6).
У цих водоростей статевий диморфізм і роздільностатевість проявляється у поколінь, що утворюють гамети (жіночий гаметоспорофіт та чоловічий гаметофіт - нанадрій). Крім того, у поколінь, що утворюють спори (жіночий гаметоспорофіт та чоловічий спорофіт), і у зооспор, що утворюються з зиготи, наявна роздільностатевість без статевого диморфізму.
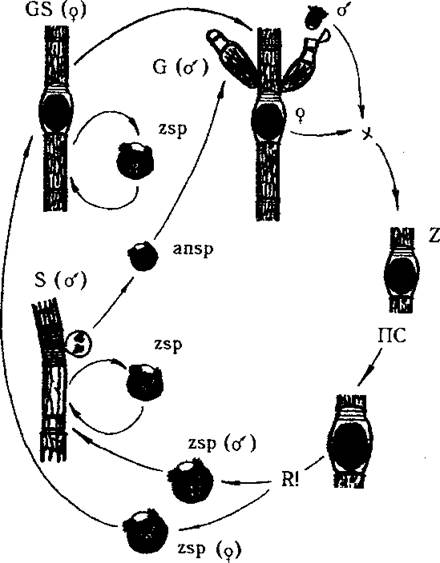
Рис. 8.6. Життєвий цикл у нанандрієвих видів роду Oedogonium: на ниткоподібному жіночому гаметоспорофіті (GS) утворюються зоооспори (zsp) та оогонії; на чоловічих спорофітах (S) - зооспори та особливі чоловічі спори - андроспори (ansp). Андроспори після виходу зі спорангію осідають на жіночій гаметоспорофіт поблизу оогонія та проростають у карликовий чоловічій гаметофіт - нанандрій (G). У верхівковій клітині нанандрію утворюються два сперматозоїди, один з яких через розрив оболонки оогонію проникає до яйцеклітини і запліднює її. Зигота (Z) переходить у стан спокою. По закінченню періоду спокою у зиготі відбувається мейоз, формуються чотири зооспори, дві з яких проростають надалі у жіночій гаметоспорофіт, а дві інші - у чоловічий спорофіт.
Різноманітність життєвих циклів
Різноманітність типів статевого розмноження, редукційного поділу, чергувань поколінь, змін форм розвитку, явища статевого диморфізму, численні випадки партеногенезу обумовлюють велику різноманітність життєвих циклів у водоростей. Ускладнення життєвих циклів відігравало суттєву роль в еволюції водоростей на рівні таксонів середнього та низького рангу, причому у різних відділах основні напрямки розвитку життєвих циклів, як правило, були різні.
Так, у деяких відділів водоростей життєві цикли досить одноманітні. Наприклад, у Rhaphidophyta, Eustigmatophyta, Dictyochophyta, Glaucocystophyta статеве розмноження не виявлено, і життєвий цикл являє собою цикломорфоз, хоча й часом досить складний.
Серед еугамних форм у кількох відділів життєвий цикл проходить без чергування поколінь і з фіксованим типом редукції кількості хромосом. Так, у еугамних Euglenophyta, Xanthophyta, Chrysophyta редукція зиготична і життєвий цикл гаплофазний. Особливість життєвого цикл у останнього відділу полягає у тому, що ядра гамет після копуляції одразу не зливаються, і замість типової зиготи утворюється особлива двоядерна зиготична циста, яка переходить у стан спокою.
Злиття ядер відбувається лише по закінченні періоду спокою. В ядрі щойно утвореної зиготи одразу відбувається мейоз. У Bacillariophyta життєвий цикл завжди диплофазний, з гаметичною редукцією і особливою стадією, що отримала назву ауксоспори.
У Dinophyta цікавою особливістю циклу розвитку є те, що зигота навіть у одного й того ж виду може проростати як з редукційним поділом, так і без нього, тобто тип редукції жорстко не фіксований, на відміну від представників інших відділів.
Складні цикли з чергуванням двох поколінь зустрічаються лише у чотирьох відділах водоростей - Phaeophyta, Haptophyta, Chlorophyta та примітивних Rhodophyta. У продвинутих в еволюційному відношенні червоних водоростей з класу Флоридей у життєвому циклі чергуються три покоління - спорофіт, гаметофіт та карпоспорофіт. Цікаво, що, за вийнятком Haptophyta, всі ці відділи містять багато складних за морфологічною будовою видів.