Альгологія - Костіков І.Ю. - 2009-2013
Розділ 17. Діатомові водорості – Bacillariophyta
Відділ діатомових водоростей нараховує більше 20 тис. видів1, і за різноманітністю знаходиться на другому місці після зелених водоростей. Діатомові водорості виникли порівняно недавно - біля 130 млн років тому, у крейдяний період фанерозою, майже одночасно з квітковими рослинами. У теперішній час діатомові перебувають у стані біологічного прогресу і є домінуючою групою у морських та прісноводних біотопах.
Всі діатомові водорості - це кокоїдні одноклітинні або колоніальні організми, розміри яких коливаються в діапазоні 4-2000 мкм при середніх значеннях 20-50 мкм.
Діатомові водорості належать до трубчастокристних організмів з філи страменопілів, мають вторинно симбіотичні пластиди, що походять від родопластів. Найяскравіша специфічна ознака діатомових водоростей - це наявність кремнеземового панциру.
Систематичні ознаки відділу
Пігменти та запасні поживні речовини
Фотосинтетичні пігменти у діатомових водоростей представлені хлорофілами а та с, β- та ε- каротинами. З ксантофілів переважає жовтий пігмент фукоксантин, який обумовлює коричневе забарвлення пластид; у менших кількостях представлені діатоксантин, діадіноксантин та неоксантин.
Основним продуктом асиміляції є хризоламінарин, додатковими - олія та волютин. Продукти асиміляції відкладаються або безпосередньо у цитоплазмі, або у вакуолях.
Цитологічні ознаки
Клітинні покриви у діатомових водоростей високо специфічні і представлені мінеральним панциром, що розташовується назовні від плазмалеми. За хімічним складом панцир діатомей близький до опалу, його основу складає кремнезем (SiO2 * nH2O). Крім того, до складу панциру входять солі алюмінію, заліза, магнію, а також невелика кількість білків. Питома вага панциру в середньому складає 2,07. Цікаво, що при концентрації кремнезему в оточуючому середовищі нижчій від 0,5 г/л клітини не здатні збудувати панцир, і тому швидко гинуть. Крім того, деякі метали (зокрема, германій) здатні пригнічувати утворення панциру.
Панцир у всіх діатомових водоростей утворений двома половинами - більшою - епітекою, та меншою - гіпотекою (рис. 17.1). Кожна половина панциру складається зі стулки (вальви) та пояскового обідка (цингулюма). Краї стулки зігнуті під більш-менш прямим кутом, і утворюють зону, яка називається загином стулки. З краєм стулки у цій зоні одним боком щільно з’єднується поясковий обідок. Поясковий обідок епітеки щільно насувається на поясковий обідок гіпотеки, завдяки цьому панцир має вигляд коробки (гіпотеки), яка накрита кришкою (епітекою). Зона перекривання пояскових обідків епітеки та гіпотеки називається пояском. Відповідно, панцир діатомових можна розглядати у двох позиціях - зі стулки та з пояска. Панцир має досить складну структуру, специфічну для кожного роду та виду діатомових водоростей.
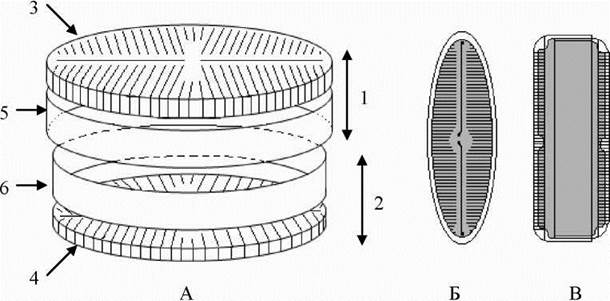
Рис. 17.1. Схема будови панцира діатомових водоростей. А - основні структурні елементи панцира; Б, В - вигляд панцира у різних площинах (Б - зі стулки; В - з пояска). 1 - епітека; 2 - гіпотека; 3 - стулка епітеки - епівальва; 4 - стулка гіпотеки - гіповальва; 5, 6 - пояскові обідки (5 - епіцингулюм, 6 - гіпоцингулюм).
Форма панциру у діатомових водоростей досить різноманітна, і характеризується, в першу чергу, типом симетрії стулки. Якщо через стулку можна провести багато вісей симетрії, таку стулку називають радіально- симетричною, або актиноморфною (рис. 17.2). Коли ж через стулку можна провести лише дві, одну або жодної вісі симетрії, стулку називають зигоморфною. Зигоморфні стулки бувають симетричними як у поздовжньому, так і поперечному напрямку (бісиметричні), симетричними лише по одній вісі - або поздовжній, або поперечній (моносиметричні стулки), дзеркально симетричними (S-подібними) або асиметричними. При вигляді з пояску панцири діатомових зазвичай (але не зважди) більш-менш прямокутні.
Стулка та її основні структурні елементи. Стулка має гомогенні ділянки, які називають гіаліновими, систему перфорацій, через яку відбувається взаємодія протопласту із зовнішнім середовищем (ареоли, шви і т.п.), систему внутрішніх та зовнішніх виростів та потовщень панциру (ребра, вирости і т.п.).
Поля. Великі гіалінові ділянки називаються полями. Якщо поле розташовується в центрі стулки, його називають центральним. Поле, яке проходить від одного полюсу стулки до протилежного, називать осьовим.
Перфорації. Найбільш поширеним типом перфорацій є ареоли. Ареола - це дрібний отвір у панцирі, закритий з одного боку тонкою перфорованою кремнеземовою платівкою - велумом. Велум може розміщуватись або на зовнішньому, або на внутрішньому боці стулки. Зрідка велум закриває ареолу з двох боків. Відкритий бік ареоли, позбавлений велуму, називається фораменом. При оптичній мікроскопії ареоли мають вигляд дрібних крапок. Якщо такі крапки розташовуються щільними правильними рядами, то система крапок набуває вигляду штриха. По відношенню до краю стулки штрихи поділяють на паралельні, радиальні та конвергентні (рис. 17.2).
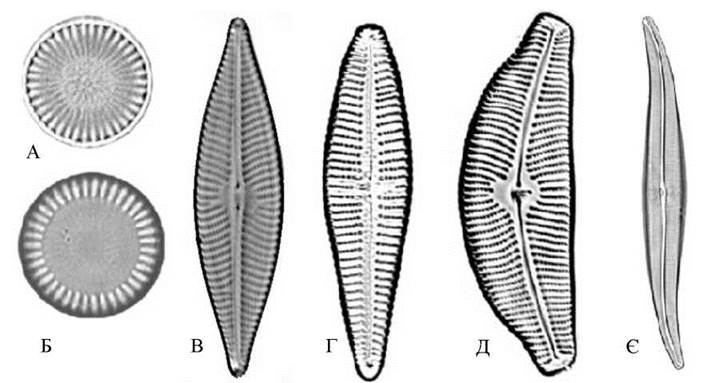
Рис. 17.2. Типи стулок діатомових водоростей: А, Б - актиноморфні; В - бісиметричні; Г, Д - моносиметричні; Є - дзеркально-симетричні (А - Cyclostephanos dubius, Б - Cyclotella meneghiniana, В - Navicula trivialis, Г - Gomphonema affine, Д - Cymbella tumida, Є - Gyrosigma acuminatum).
У водоростей з зигоморфним панциром досить часто наявний шов. У шві циркулює цитоплазма, і завдяки цьому ті діатомові водорості, які мають шов, здатні до активного ковзаючого руху. Шви поділяють на щілиноподібні та каналоподібні.
Щілиноподібний шов в оптичний мікроскоп має вигляд чіткої лінії, яка починається на полюсах клітини, проходить по осьовому полю і переривається у центральній частині стулки - у зоні центрального поля. Ділянка шва від полюса до центрального поля називається гілкою шва, тобто щілиноподібний шов має дві гілки. В товщі стулки в зоні центрального поля кожна гілка шва закінчується вертикальним каналом. Канали обох гілок, в свою чергу, з'єднуються одним горизонтальним каналом. Вся ця система з двох вертикальних та одного з'єднувального горизонтального канала розташовується у потовщенні стулки, яке називається центральним вузликом. На полюсах зовнішня щілина шва розширюється та вигинається, утворюючи полярну щілину. Потовщення стулки, в якому розташовується полярна щілина, називається полярним вузликом.
Каналоподібний шов має вигляд суцільної лінії, що не перериваючись проходить від одного полюсу клітини до іншого. Шов цього типу складається з вузької зовнішньої щілини та трубчастого каналу, який є аналогом внутрішньої щілини щілиноподібного шва. Трубчастий канал відкривається у порожнину клітини добре помітними отворами - фібулами. На відміну від щілиноподібного шва, який розміщується найчастіше по вісі стулки, каналоподібний шов розташовується, як правило, по краю стулки, у її внутрішньому потовщенні - кілі, або у зовнішньому вирості - крилі.
Потовщення стулки. У значної кількості діатомових водоростей стулка має систему потовщений ділянок, що надають поверхні стулки вигляду пральної дошки. Такі потовщення називають ребрами. При вигляді зі стулки ребра дещо нагадують штрихи, проте відмінність ребер від перфорацій стає добре помітною при розгляді панциру з пояска.
Крім ребер, стулка може мати і інші потовщення, наприклад: трубчасті вирости з опорами - фултопортули, двогубі вирости - римопортули, короткі шипи та довгі щетинки і т.п.
Пояскові обідки панциру завжди парні - один обідок (епіцингулюм) з'єднується з загином стулки епітеки, другий (гіпоцингулюм) - з загином стулки гіпотеки. Поясковий обідок епітеки протилежним від загину стулки боком завжди насувається на поясковий обідок гіпотеки, утворюючи зону з’єднання епітеки та гіпотеки - поясок.
Поясковий обідок кожної половини панциру може бути простим або складним. Простий обідок складається лише з однієї замкненої у кільце стрічки. Цей тип обідка у діатомових водоростей є найпоширенішим. Складні обідки зустрічаються у значно меншої кількості видів. Вони утворені кількома послідовно вкладеними одна в одну кільцеподібними стрічками. Кільце, яке безпосередньо прилягає до стулки, називають стулковим кільцем, або вальвокопулой. Кільце, з'єднується з вальвокопулою та всі наступні послідовно приєднані кільця пояскового обідка називають вставними обідками. Вставні обідки половин панциру, за допомогою яких епітека з'єднується з гіпотекою, називають з'єднувальними.
Вставні обідки найчастіше суцільні, рідше кожний обідок утворений двома з'єднаними між собою напівкільцями або складаються з числених щільно з'єднаних між собою сегментів.
У деяких діатомових водоростей поясковий обідок може розростатися в порожнину клітини, утворюючи розташовану паралельно до стулки неповну перегородку - септу.
До внутрішнього боку панциру щільно притиснута плазмалема.
Чимало видів діатомових водоростей здатні виділяти слиз. За допомогою слизу водорості утворюють слизові трубки (наприклад, Cymbella), різноманітні слизові ніжки (Rhoicosphenia, Gomphonema), з'єднуються у колонії (Tabellaria, Fragilaria). У водоростей, які мають шов, слиз відіграє надзвичайну роль у процесі активного ковзаючого руху клітини. Крім того, за допомогою слизу під час статевого процесу клітини зближуються і утримуються разом доти, доки відбувається злиття їх цитоплазм - плазмогамія.
Ядерний апарат діатомових типово евкаріотичний. В інтерфазі ядро завжди одне, розташовується в центрі клітини. Як і у інших водоростей-страменопілів, ядро структурно зв'язано з оболонкою хлоропластів, оскільки зовнішня мембрана ядра переходить у зовнішню мембрану хлоропластної ендоплазматичної сітки. Між мембранами ядерної оболонки можуть утворюватись мікротубулярні частини тричленних джгутикових мастигонем (ретронем), але через відсутність у багатьох діатомей монадних стадій цей процес вдається спостерігати зрідка. Біля поверхні ядра, як правило, розташовується 1-2 комплекси Гольджі.
Мітоз у діатомових водоростей відкритий, центріолі відсутні. Замість них в інтерфазі на латеральному боці ядра майже біля пояска є щільно притиснуте до ядерної оболонки дрібне тіло, яке і виконує функцію центру організації мікротрубочок - МТОЦ.
В профазі біля МТОЦ з'являється дрібне шарувате тіло, за допомогою якої, ймовірно, МТОЦ розділяється на дві субодиниці, причому кожна субодиниця набуває форми диску. Далі між дисками утворюються мікротрубочки веретена, при цьому самі диски розходяться на протилежні полюси ядра і розташовується біля поверхні стулки: один - під епітекою, другий - під гіпотекою. У пізній профазі ядерна оболонка руйнується, мікротрубочки веретена переміщуються у ядерну зону.
У метафазі веретно диференціюється на так зване центральне та радіальне. Центральне веретено проходить через центр ядерної зони і з'єднує обидва диски. Мікротрубочки центрального веретена численні, розташовуються по колу і утворюють два перекриваючих напівциліндри. Термінальною частиною кожний напівциліндр приєднаний до диску. Радиальне веретено складається з поодиноких відносно нечисленних мікротрубочок, які одним кінцем прикріплюються до диску, протилежним - до хромосоми.
В анафазі мікротрубочки центрального веретена подовжуються в зоні перекривання напівциліндрів, розсуваючи диски з мікротрубочками радіального веретена до протилежних стулок. Разом з мікротрубочками радіального веретена до протилежних стулок розходяться і хромосоми.
У телофазі центральне веретено руйнується, хромосоми від'єднуються від ниток радіального веретена і ядерна оболонка відновлюється. Плазмалема у площині паралельній до стулок утворює борозну поділу, яка розділяє материнську клітину на дві дочірні, при чому кожна клітина отримує по одному диску з пучком звільнених від хромосом радіальних мікротрубочок.
Далі кожний полярний диск разом з мікротрубочками мігрує на протилежний бік ядра - до поверхні майбутньої нової стулки. В цей час комплекс Гольджі та ендоплазматична сітка починають відчленовувати велику кількість мембранних пухирців, заповнених кремнеземом. Ці пухирці мігрують до плазмалеми, під якою починають зливатися між собою, утворюючи силікалему - велику одномембранну цистерну, в якій відбувається формування нової стулки. Рухом та впорядкованим злиттям кремнезем-вмісних пухирців з силікалемою "керують" мікротрубочки, які залишились від радіального веретена.
Таким чином, у діатомових водоростей процес утворення кремнеземового панциру тісно пов'язаний з ядерним апаратом, зокрема, з центром організації мікротрубочок. Цікавим є й те, що поділу ядра завжди передує поділ хлоропластів, який відбувається переважно у пізній інтерфазі.
Фотосинтетичний апарат діатомових - це вторинно-симбіотичними пластиди родофітного типу. Хлоропласти переважно займають пристінне положення, їх кількість і форма специфічні для різних родів. Залежно від освітлення, хлоропласти здатні дещо змінювати положення - переміщуватись вглиб клітини або підійматися до її поверхні. Здатність до руху обумовлена наявністю скоротливих мікрофібрил, що з'єднують зовнішню мембрану пластиди з плазмалемою.
Хлоропластна оболонка має чотири мембрани, з яких дві зовнішні утворюють хлоропластну ендоплазматичну сітку, що переходить у ядерну оболонку. Між зовнішніми та внутрішніми мембранами хлоропласта є перипластидний простір (рис.).
Тилакоїди по три зібрані у ламели; безпосередньо під оболонкою хлоропласту розташовується оперезуюча ламела. Досить часто в хлоропласті є голий піреноїд, який, на відміну від багатьох інших водоростей-страменопілів, може бути пронизаний парами тилакоїдів. На полюсах пластид компактно розташовується замкнена у кільце хлоропластна ДНК, яка містить біля 120-130 тис. пар нуклеотидів.
Мітохондрій зазвичай кілька. Вони розміщуються у периферичній частині клітин і мають лише трубчасті кристи.
Вакуолярний апарат включає 4 типи вакуолей: з клітинним соком, волютином, хризоламінарином та олією. У водоростей з актиноморфними панцирами вакуолі з клітинним соком переважно дрібні і численні. Якщо панцир зигоморфний, то найчастіше таких вакуолей дві. Вони розміщуються по поздовжній вісі клітини, відтисняючи цитоплазму до периферії, проте в центрі клітини залишається цитоплазматичний місток, в якому розташовується ядро. Вакуолі з волютином - тільця Бючлі - є резервним депо поліфосфатів. Вони забарвлені у слабкий блакитний колір, і їх положення є специфічним для різних родин діатомових водоростей.
Джгутиковий апарат. Монадні стадії представлені виключно голими одноджгутиковими сперматозоїдами, і відомі лише у кількох родів діатомових водоростей. Джгутик вкритий ретронемами, що утворюються в хлоропластному ендоплазматичному ретикулюмі та міжмембранному просторі ядерної оболонки. Аксонема нетипова, оскільки представлена лише дев'ятьма дублетами периферичних мікротрубочок, а дві центральні мікротрубочки відсутні (формула 9+0). Є відміни і в будові базального тіла - замість триплетів воно утворено дублетами мікротрубочок, і, подібно до диктиохофітових водоростей (див. розділ 18), притиснуто до ядерної мембрани.
Рух. У діатомей виявлено три принципово різних типи рухів: а) за допомогою джгутиків, який здійснюється лише у сперматозоїдів у деяких центричних діатомей; б) амебоїдні рухи протопластів гамет під час статевого процесу; в) ковзаючий рух у вегетативних клітин видів, які мають розвинені шви. Механізми руху перших двох типів аналогічні тим, які наявні у багатьох інших одноклітинних водоростей, найпростіших та грибоподібних організмів. Механізм ковзаючого руху є унікальним.
Ковзаючий рух вегетативних клітин відбувається за участю слизових фібрил, які секретуються через зовнішню щілину шва. При цьому одним кінцем слизова фібрила прикріплюється до субстрату, а протилежним залишається з'єднаною з вбудованими у плазмалему у зоні шва трансмембранними структурами. Трансмембранні структури включають білок та АТФазу, і, в свою чергу, асоційовані з пучками скоротливих актинових мікрофібрил, що розташовуються під плазмалеммою вздовж шва. Біля середньої частини гілки шва біля трансмембранних структур розташовуються субмікроскопічні пухирці з фібрилярним матеріалом полісахаридної природи, які називають кристалоїдними тільцями.
Під час руху вміст кристалоїдних тілець виштовхується у щілину шва поблизу центрального вузлика, ущільнюється та структурується, перетворюючись на слизову фібрилу з кислих мукополісахаридів. Для прикріплення слизової фібрили до субстрату необхідним є кальцій. За допомогою скоротливих мікрофібрил трансмембранні структури з прикріпленими до них слизовими фібрилами протягуються по плазмалемі вздовж шва. Як наслідок, клітина рухається у напрямку, протилежному до напрямку переміщення слизової фібрили. Коли фібрила досягає полярної щілини, вона відривається від плазмалеми і залишається на субстраті. За допомогою метиленового синього за положенням відокремлених слизових фібрил, що залишилися на субстраті, на великих збільшеннях мікроскопу можна реконструювати треки руху клітини.
Форми треків відповідають формі шва: у водоростей з прямим швом (наприклад, Navicula, Pinnularia) треки прямі; з вигнутим (Nitzschia) - широкі дугоподібні, з S-подібним швом (Gyrosigma) - сигмовидні. У одношовних водоростей з увігнутими стулками (Cocconeis) треки дрібнохвилясті. Специфічні рухи здійснюють шовні водорості, прикріплені до субстрату слизовими ніжками: наприклад, Gomphonema періодично стрімко обертається навколо ніжки.
Напрямок руху може бути обумовленим як фототаксисом, так і хемотаксисом, хоча безпосередні фото- та хеморецептори залишаються не встановленими. Рух у діатомей реверсивний: в процесі руху слизові фібрили почергово секретуються різними гілками шва, внаслідок чого клітина рухається вперед та назад, при цьому трек в одному з напрямків є довшим за інший.
Типи морфологічних структур тіла
Діатомові водорості мають виключно кокоїдний тип морфологічної структури тіла, хоча в деяких випадках ниткоподібні колонії діатомей можуть бути помилково ідентифіковані як нитчасті форми (наприклад, Melosira).
Розмноження та життєві цикли
Представники відділу розмножуються вегетативним поділом клітин надвоє та статевим шляхом. Всі діатомеї мають єдиний тип життєвого циклу - диплофазний цикл з гаметичною редукцією без зміни поколінь та зі специфічною ростовою стадією - ауксоспорою.
Вегетативний поділ надвоє є основним типом розмноження і притаманний всім видам діатомових. Перед поділом клітина за рахунок накопичення олії або активного поглинання води дещо набрякає. Внаслідок цього починається розсування епітеки та гіпотеки. Далі діляться хлоропласти, відбувається мітоз. Після мітозу кожна дочірня клітина отримує одну половину материнського панциру, і далі з силікалеми будує другу половину. Материнська половина завжди стає епітекою дочірньої клітини, оскільки нова стулка, що формується у силікалемі, обмежена краями пояскового обідка панциру, що був отриманий від материнської клітини.
По завершенні утворення стулки, кремній-вмісні пухирці переміщуються у зону, розташовану під поясковим обідком материнської половини панциру, і утворюють вальвокопулу гіпотеки. Вальвокопула з'єднується зі стулкою гіпотеки, і формування невистачаючої половини панциру завершується.
Таким чином, внаслідок вегетативного поділу надвоє клітина, що отримала у спадок материнську епітеку, зберігає розміри материнської клітини. У другої клітини половина панциру, що у материнської клітини була гіпотекою, стає епітекою. Тобто розмір другої клітини зменшується. Оскільки кремнезем не здатний розтягуватись, то в принципі розміри клітин у популяції мали б швидко зменшуватись. Проте в дійсності такого не відбувається, оскільки у діатомових водоростей існують певні механізми підтримання сталих розмірів. Серед них до провідних належать механізми відтворення оптимальних розмірів через стадію ауксоспори, переважно внаслідок статевого процесу.
Ауксоспора (або спора, що росте) - це особлива клітина, яка здатна до росту. Клітина, яка перетворюється на ауксоспору, скидає панцир, і починає швидко збільшуватись у розмірах. По закінченні процесу росту протопласт ауксоспори утворює епітеку та гіпотеку, формуючи т.з. ініціальну клітину, розміри якої зазвичай наближуються до максимально можливих для даного виду. Хоча ауксоспори можуть утворюватися внаслідок різних процесів (наприклад, при досягненні клітинами мінімальних можливих розмірів або при пошкодженнях панциру), проте найчастіше вони виникають після статевого процесу - з зиготи, що проростає.
Статевий процес. При статевому процесі дві клітини вкриваються спільним слизом, протопласти збільшуються у розмірах і розсувають половинки панцирів. Подальший розвиток у різних видів відбувається різними шляхами, обумовлюючи прояв певного типу статевого процесу. Проте спільним в усіх випадках є те, що ядра кожної з клітин від одного до кількох разів діляться, причому один з поділів обов'язково є редукційним.
У водоростей з актиноморфними стулками найчастіше зустрічається оогамний статевий процес. У водоростей з зигоморфними стулками найбільш поширеними типами статевого процесу є ізо- та гетерогамія. Внаслідок ізогамії утворюються дві зиготи, а гетерогамії - або дві, або одна.
При оогамії в одній з клітин - оогонії, відбувається мейоз, і три з чотирьох гаплоїдних ядер дегенерують. Протопласт з останнім, життєздатним ядром перетворюється на одноядерну гаплоїдну яйцеклітину. В чоловічій клітині - антеридії, який часто називають сперматогонієм, відбувається від одного до кількох поділів, причому останнім з них є мейоз. Як наслідок, в сперматогонії утворюється від двох до багатьох дрібних чоловічих гамет. У різних видів ці гамети можуть мати рухливі або нерухомі джгутики, або бути позбавленими джгутиків взагалі. Рухливі чоловічі гамети називають сперматозоїдами, нерухомі - спермаціями. Сперматозоїд або спермацій запліднює яйцеклітину. Утворюється зигота, яка без періоду спокою проростає в ауксоспору.
При ізогамії кожна материнська клітина ділиться мейотично, утворюючи чотири гаплоїдних ядра, два з яких дегенерують. Далі протопласт гаметангія ділиться без поділу ядер, і як наслідок, в кожному гаметангії утворюється по дві одноядерні нерухомі гамети. Копуляція відбувається між парами гамет з різних гаметангіїв. Оскільки гаметангії при цьому утримуються поруч спільним слизом і стикаються поясками, для забезпечення фізичного контакту між гаметами виявляється достатнім лише деяке збільшення протопластів гамет у розмірі. Після копуляції кожна пара гамет утворює зиготу, яка перетворюється на ауксоспору.
Гетерогамія з утворенням двох зигот відбувається майже так само, як і ізогамія, за винятком того, що гамети з одного гаметангію, який вважається чоловічим, рухливі, а другого - нерухомі. Чоловічі гамети за рахунок амебоїдних рухів протопласту переміщуються до жіночих, зливаються з ними, і утворюють також дві зиготи, що проростають в ауксоспори.
При гетерогамії з утворенням однієї зиготи протопласти майбутніх гаметангіїв діляться мейотично. В кожному гаметангії три з чотирьох гаплоїдних ядер дегенерують, і утворюється лише одна гамета. Копуляція відбувається внаслідок злиття рухливої гамети з чоловічого гаметангія з нерухомою жіночою гаметою. Як наслідок, уговорюється лише одна зигота, і, відповідно, лише одна ауксоспора.
Особливим способом статевого процесу у діатомей є автогамія. У такому процесі бере участь лише одна клітина. Її ядро редукційно ділиться, далі два гаплоїдних ядра дегенерують, а два інших зливаються, відновлюючи диплоїдне ядро і утворюючи зиготу, що перетворюється на ауксоспору.
Таким чином, статевий процес у діатомових не лише забезпечує певний рівень рекомбінантної мінливості, але й виконує функцію підтримання певного діапазону розмірів клітин у популяції, протидіючи тим самим можливому виродженню популяцій внаслідок зменшення клітин в процесі їх вегетативного поділу.
Життєвий цикл у діатомових водоростей диплофазний, з гаметичною редукцією, без чергування поколінь та з унікальною стадією - ауксоспорою.
Спочиваючі стадії представлені двостулковими кремнеземовими цистами і відомі лише у деяких морських центричних діатомей. Утворення цисти починається з поділу протопласту клітини. Далі материнський панцир скидається, кожний дочірній протопласт вкриваєтеся двома опуклими стулками, при цьому пояскові обідки не розвиваються. Стулка, яка утворилась першою, несе на поверхні різноманітні потовщення (гребні, шипи, шипики). Друга стулка залишається гладкою.
Система відділу
Провідні ознаки, за якими будується система Bacillariophyta, пов'язані з будовою кремнеземового панциру. З цим комплексом ознак корелює чимало ознак, пов'язаних з будовою протопласту та особливостями статевого процесу. Сьогодні існують дві основні групи систем діатомових водоростей: класичні морфологічні та морфолого-цитологічні.
Принципи класичної морфологічної системи розроблені у ХІХ ст. німецьким альгологом Ф. Густедтом, який запропонував виділяти таксони Bacillariophyta за помітними в оптичний мікроскоп морфологічними особливостями панциру. У ХХ ст. різні автори неодноразово пропонували модифікації системи Густедта. Наприклад, в Україні широко використовується модифікація, запропонована О.В. Топачевським та О.П. Оксіюк. У країнах Західної Європи та Америки однією з найпопулярніших є варіант системи Густедта, розроблений Х. Ланже-Берталоттом та К. Краммером.
Морфолого-цитологічна система була запропонована у другій половині ХХ ст. британськими альгологами Ф. Раундом, Р. Крауфордом та Д. Манном на основі даних, отриманих при вивченні ультратонкої будови панциру за допомогою скануючого електронного мікроскопу.
Наприкінці ХХ ст. було здійснено перші спроби тестування систем обох типів за допомогою методів молекулярної таксономії. Проте дерева, побудовані за результатами секвенування гену, що кодує 18S рибосомальну РНК показали, що ані класичні, ані морфолого-цитологічні системи не мають достатнього узгодження з молекулярними дендритами.
Таким чином, питання про систему, що адекватно відображує філогенію відділу, залишається відкритим. Нижче матеріал подано за морфолого- цитологічною системою Ф. Раунда, Р. Крауфорда та Д. Манна, яка дещо краще узгоджується з молекулярними даними, ніж варіанти класичної системи Густедта.
За цією системою Bacillariophyta включає три класи, які виділяються за симетрією стулок та наявністю шва: Coscinodiscophyceae, Fragilariophyceae та Bacillariophyceae.
Клас Coscinodiscophyceae (Centrophyceae)
Об'єднує водорості з актиноморфними стулками, які завжди позбавлені шва. У більшості випадків стулки косцинодискофіцієвих круглі, радіально- симметричні, через що представників класу часто називають центричними діатомеями. За палеоальгологічними даними саме від водоростей цього класу походять інші групи діатомей. Лише у центричних діатомей виявлено оогамні статеві процеси та рухливі сперматозоїди. Представники цього класу мешкають переважно у планктоні морів та прісних водойм. Найбільш відомі представники цього класу належать, в першу чергу, до порядків Thalassiosirales та Melosirales.
Thalassiosirales включає водорості, у яких на стулці є фултопортули і ареоли закриті велумом з внутрішнього боку. Системи ареол утворюють тангентальні або радіальні ряди. Пластиди численні, дископодібні. Вважають, що наявність внутрішнього велуму є ознакою примітивності цього порядку, оскільки такий тип велуму відомий у найдавніших викопних діатомей з вимерлого порядку Pyxidisculales.
Представники Thalassiosirales вельми поширені у планктоні морів та континентальних водойм. Зокрема, в морях (в тому числі, Чорному та Азовському) одним з домінантів є Thalassiosira. Клітини талассіозіри поодинокі або з'єднані у слизом у нетривкі колонії. Стулки круглі, з ареолами, що утворюють тангентальні штрихи.
До іншого характерного роду порядку - Cyclotella - належать переважно прісноводні планктонні види. Клітини циклотели найчастіше поодинокі; стулки, подібно до талассіозіри, круглі, але ареоли утворюють не тангентальні, а радіальні штрихи, які особливо чітко помітні біля краю стулки.
Melosirales вважається еволюційно більш продвинутим порядком: фултопортули у них відсутні, велум розташовується на зовнішньому боці ареол. Останні переважно не утворюють чітких правильних штрихів. Пластиди у мелозиральних численні, дрібні, лопатеві, розміщуються у периферичному шарі цитоплазми.
Типовий представник порядку - Melosira - надзвичайно характерний для планктонних угруповань прісних водойм. Панцири мелозіри мають високі загини стулки та складні пояскові обідки, через що в препаратах клітини лежать переважно з пояска і мають видовжено-циліндричну форму. На стулці розташовується кілька римопортул, дуже дрібні ареоли, інколи - кільце дрібних шипів. Через римопортули виділяється слиз, за допомогою якого дочірні клітини після поділу залишаються з'єднаними стулками, утворюючи пари та триплети клітин. Пари та триплети, залишаючись разом, обумовлюють формування типових для цієї водорості ниткоподібних колоній.
У Melosira досить детально досліджено статевий процес, який представлений оогамією. На початку статевого процесу в одній з клітин відбувається мейоз, при цьому після першого поділу (мейоз 1) одне з ядер дегенерує, і не переходить у фазу мейозу 2; друге, життєздатне ядро, ділиться, і знов одне з ядер дегенерує. Цитокінез при поділах мейоз 1 та 2 не відбувається. Таким чином, у жіночому гаметангії утворюється одна яйцеклітина.
В клітині, що виконує функцію чоловічого гаметангію, протопласт набрякає, обумовлюючи скидання панциру. Далі на поверхні протопласту з'являються чотири джгутики, відбувається мейоз і утворюється чотириядерна гола сперматогенна клітина, що зберігає всі чотири джгутики. Далі одночасно від сперматогенної клітини відбруньковуються чотири одноджгуткові одноядерні сперматозоїди. Брунькування відбувається таким чином, що в гамети не потрапляють пластиди: вони залишаються в центрі сперматогенної клітини, утворюючи т.з. залишкову клітину, що позбавлена ядра і незабаром руйнується. Далі сперматозоїди запліднюють яйцеклітину, яка перетворюється на ауксоспору.
Клас Fragilariophyceae
Об'єднує водорості з зигоморфними стулками, які позбавлені швів, але мають осьове поле. На стулці фрагілярієфіцієвих, як правило, розташовується одна римопортула, яка вважається попередником шва. В межах цього класу, на відміну від Coscinodiscophyceae, повністю відсутні джгутикові стадії. Найбільш поширені представники класу належать до порядків Fragilariales, Tabellariales та Licmophorales, що відрізняються, в першу чергу, положенням римопортули (полярна чи зміщена до центру) та наявністю вставних обідків із септами.
Для Fragilariales характерними є наявність на полюсі стулки однієї римопортули, розміщеної під кутом до поздовжньої вісі; вставні обідки відсутні або наявні, проте в останньому випадку позбавлені септ. Клітини містять від двох великих до багатьох дрібних хлоропластів, розміщеих переважно в периферичному шарі цитоплазми під стулкою. Більшість представників цього порядку утворює колонії, за формою яких водорості можна розпізнавати на рівні роду навіть без виготовлення постійних препаратів панцирів.
Зокрема, клітини Fragilaria є поодинокими або утворюють колонії у вигляді стрічок, в яких панцири з'єднуються стулками. При вигляді з пояска окремий панцир має видовжено-прямокутну форму; зі стулки панцир видовжений білатерально-симетричний, на полюсах більш-менш дзьобоподібний. У постійних препаратах, виготовлених з очищених від цитоплазми панцирів, на стулці добре розрізняються осьове поле та чергово розміщені штрихи. Види роду є переважно мешканцями перифітонних та бентосних угрупувань прісних водойм.
За будовою панциру вельми схожа с фрагілярією прісноводна планктонна водорість Asterionella, яка, проте, добре відрізняється за будовою колоній. В них клітини з'єднані у зірчасті колонії за допомогою слизу та дрібних шипіків, розташованих на полюсі стулки.
Рід Diatoma у живому стані розрізняється за наявністю зигзаг-подібних колоній. В них з'єднання клітин також здійснюється за допомогою слизу та полярних шипиків на стулці. Крім того, яскравою ознакою цього роду є наявність на внутрішньому боці стулки грубих поперечних ребер, які надають клітині характерної поперечної посмугованості. Діатому можна часто зустріти у планктонних та бентосних угрупуваннях прісних водойм.
Порядок Tabellariales відрізняється від попереднього тим, що римопортула розташовується біля центру стулки та наявністю вставних обідків з септою, що перекриває майже половину такого обідка. Як наслідок, система септ розділяє панцир на кілька напіввідкритих камер. В кожній такій камері знаходиться одна стічкоподібна пластида. Тому з пояска одна клітина табелляріальної водорості нагадує колонію з кілької клітин з простими поясками, з'єднаних стулками у вигляді невеличкого паркану.
Види типового роду Tabellaria дуже поширені у бентосних та перифітонних угрупуваннях прісних водойм. Зазвичай клітини табеллярії з'єднуються між собою кутами за допомогою слизу та шипиків на полюсах стулки, утворюючи зигзаг-подібні колонії, схожі на такі у діатоми.
Порядок Licmophorales включає переважно морські види перифітонних та бентосних діатомей, які мають клиновидні при вигляді з пояска панцири, що прикріплюються до субстрату за допомогою слизових ніжок. Римопортула у цих водоростей одна, розташовується біля полюсу клітини. Типовий рід порядку - Licmophora - належить до морських водоростей-космополітів і зустрічається практично всюди в обростаннях каміння, черепашов молюсків, таломів водоростей-макрофітів та морських вищих рослин.
Клас Bacillariophyceae
До цього класу включені водорості з зигоморфними стулками, у яких на одній або обох стулках є шов. Згідно з сучасними поглядами на еволюцію діатомових водоростей, шов утворився з римопортули, що поступово видовжувалась.
Шов є структурою, з якою пов'язана здатність бацилярієфіцієвих водоростей до активного ковзаючого руху. Поява такої здатності розглядається як крупний ароморфоз. Тому ознаки, пов'язані з швом (зокрема, його тип, кількість, положення на стулці, ступінь розвитку і т.д.) покладені в основу поділу Bacillariophyceae на порядки.
Одним з наслідків переходу до рухливого способу життя стала поступова редуція здатності утворювати колонії з клітин, що залишились разом після поділу. Тому зірчасті, парканоподібні та стрічкоподібні колонії, які дуже характерні для Fragilariophyceae, в межах класу Bacillariophyceae майже не зустрічаються. Клітини бацилярієфіцієвих або поодинокі, або утворюють колонії, в яких водорості занурені у спільний колоніальний слиз, чи знаходяться у слизових трубках, або прикріплюються до субстрату слизовими ніжками.
У водоростей з Bacillariophyceae джгутикові стадії повністю відсутні, статеві процеси представлені ізо-, гетеро- та автогамією.
Bacillariophyceae поділяють на десять порядків, з яких нижче розглядаються сім (табл. 17.1): Eunotiales, Cymbellales, Achnanthales, Naviculales, Bacillariales, Rhopalodiales, Surirellales.
Таблиця 17.1. Провідні ознаки основних порядків Bacillariophyceae
Порядок |
Шов |
Симетрія стулки |
Eunotiales |
недорозвинений |
моносиметричні |
Cymbellales |
щілиноподібний, на обох стулках (на одній стулці може бути недорозвиненим) |
моносиметричні |
Achnanthales |
щілиноподібний, на одній стулці |
бісиметричні |
Naviculales |
щілиноподібний, на обох стулках |
бісиметричні, S- подібні |
Bacillariales |
каналоподібний, у кілі, по одному на стулці, фібули розрізняються добре |
моно- та бісиметричні |
Rhopalodiales |
каналоподібний, по одному на стулці, фібули не помітні |
моносиметричні |
Surirellales |
каналоподібний, у криловидному вирості, по два на стулці |
бі-, моно- та асиметричні |
До Eunotiales входять водорості з дорзовентральними стулками, симетричними по відношенню до поперечної вісі. На вентральному боці розташовується недорозвинений шов, щілини якого починаються на полюсах стулки. Хоча евноціальні водорості мають щілиноподібний шов, через його недорозвиненість вони не здатні до активного руху.
Види типового роду Eunotia населяють бентосні та перифітонні угрупування оліготрофних кислих водойм, і є характерними представниками болотної альгофлори.
Cymbellales об'єднує види, у яких на одній зі стулок є розвинений щілиноподібний шов, за допомогою якого клітини можуть активно рухатись. На протилежній стулці також є шов, який у деяких видів недорозвинений, і нагадує шов евноціальних (наприклад, Rhoicosphenia), у інших - розвинений повністю (Gomphonema, Cymbella, Encyonema). Стулки у цимбеляльних водоростей асиметричні принаймні по відношенню до однієї з вісей. Наприклад, у Rhoicosphenia та Gomphonema стулки симетричні лише по відношенню до поздовжньої вісі, через що мають обриси, наближені до булавоподібних або клиноподібних. У Cymbella та Encyonema навпаки, стулки симетричні щодо поперечної вісі, і тому є дорзовентральним.
Види з Cymbellales населяють переважно бентос та перифітон прісних водойм. Так, Rhoicosphenia часто трапляється в обростаннях вищих водних рослин. Її клітини, булавоподібні зі стулки та клиноподібні й вигнуті з пояска, прикріплюються до субстрату довгими слизовими ніжками. Цей слиз секретується через відкриті пори, розташовані на звуженому кінці стулки. Хлоропласт у ройкосфенії один, чотирилопатевий; лопаті починаються від піреноїда, притиснутого до пояска.
За обрисами клітин види роду Gomphonema дещо нагадують попередній рід - стулки в них булавоподібні, з пояска клітини клиноподібні, але, на відміну від ройкосфенії, не вигнуті. Водорості мешкають переважно у перифітоні, і прикріплюються до підводних субстратів слизовими ніжками, що утворюються завдяки секреції слизу через систему субмікроскопічних пор, розташованих на полюсі звуженого боку стулки.
Види роду Cymbella нагадують півмісяці, що знаходяться на верхівках розгалужених слизових ніжок. На вигнутому боці клітини розташовується хлоропласт з піреноїдом. Майже посередині стулки проходить шов, полярні щілини якого відігнуті на дорзальний бік. Біля центрального вузлика в оптичному мікроскопі на постійних препаратах можна спостерігати одну-кілька крапок, що представляють особливий тип перфорацій - стигми.
Рід Encyonema за формою клітин виглядає майже так само, як цимбелла. Проте клітини енционеми розташовуються переважно всередині слизових трубок, хлоропласт розміщений на вентральному боці клітини, полярні щілини шва також відігнуті на вентральний бік. Стигми у Encyonema відсутні.
Найяскравішою особливістю Achnanthales є наявність шва лише на одній зі стулок - або на епітеці, або на гіпотеці. Розташування шва лише на одній стулці пов'язано з пристосуванням цих водоростей до перифітонного способу життя. Клітини ахнантальних водоростей поодинокі, розташовуються на поверхні субстратів притискаючись до них стулкою зі швом.
В бентосних морських угрупуваннях, переважно на дрібному камінні літоралі та супраліторалі, масово розвиваються види роду Achnanthes. Зі стулки клітини цієї водорості бісиметричні, проте при розгляді клітин з пояска видно, що стулка за швом увігнута, безшовна - опукла. Переміщуючись по субстрату за допомогою шва, водорість обирає ділянки з оптимальним освітленням, і зупиняється. Далі одна з полярних щілин шва секретує слизову ніжку. Пластиди у водорості переважно численні, дископодібні.
У прісних водоймах у перифітонних та бентосних угрупуваннях масово розвивається схожі на попередній рід види роду Achnanthidium. Вони також мають бісиметричні стулки, з яких шовна стулка увігнута, а безшовна - опукла. За рахунок секреції слизу полярною щілиною, клітини утворюють ніжку, якою під час вегетації прикріплюються до субстрату. Від Achnanthes цей рід відрізняється не лише за поширенням, але й за наявністю однієї пластиди, розташованої вздовж одного з боків стулки, а також помітної на постійних препаратах підковоподібної структури біля центрального поля безшовної стулки.
В прісних водоймах найхарактернішими домінантами обростань вищих водних рослин та зелених водоростей-макрофітів є види роду Cocconeis. Їх стулки широкоеліпсоїдні, пояскові обідки низькі. Клітини Cocconeis не утворюють слизових ніжок, а міцно притискаються до субстрату шовною стулкою. Кожна клітина містить один лопатевий пристінний хлоропласт у вигляді літери "С" з одним-кількома дрібними видовженими голими піреноїдами.
Naviculales є найбагатшим за видовим складом порядком не лише в межах класу, але й відділу в цілому. водорості цього порядку мають розвинені шви на обох стулках. Самі стулки або бісиметричні, або S-подібні (дзеркально симетричні). Основними критеріями, що використовуються при поділі на родини та роди, є форма стулки, особливості будови щілин шва, структура штрихів та ребер. Додаткові ознаки, що дозволяють розрізняти роди у живому стані, пов'язані, в першу чергу, з морфологією пластид. Представники Naviculales поширені майже в усіх типах морських та прісноводних угрупувань. Характерними прикладами навікуляльних водоростей є роди Navicula, Craticula, Gyrosigma, Pinnularia.
Види одного з найчисленніших родів - Navicula - мають досить просту будову: стулки за обрисами ланцетоподібні, і нагадують човник. По середині осьового поля проходить прямий щілиноподібний шов. На поверхні стулок розташовуються паралельні, радіальні або конвергентні штрихи, утворені правильними рядами ареол. Центральне поле відносно невелике, і не доходить до країв стулки. При вигляді з пояска клітини видовжено-прямокутні. Пояскові обідки прості, і не мають проміжних обідків. Хлоропластів два, вони пластинчасті, розташовуються вздовж пояскових обідків на різних боках стулки і містять по одному паличкоподібному піреноїду.
За формою панциру, наявністю прямого шва та штрихів, утворених лише ареолами, схожим з навікулою є рід Craticula. Проте центральне поле у кратікули майже відсутнє, а ареоли розташовуються таким правильними рядами, що поверхня стулки здається порисованою двома системами штрихів - паралельних та перпендикулярних. Пластид дві, майже такі самі, як у навікули, але піреноїди не паличкоподібні, а широкоеліпсоїдні.
У видів роду Gyrosigma система перфорацій стулки схожа із кратикулою. Але стулки у гіросигми не бісиметричні, а дзеркально-симетричні, і нагадують неповну літеру "S". Центральні щілини шва або відігнуті на протилежні боки, або закінчуються коротким Т-подібним розгалуженням. Хлоропластів два; їх форма та розташування таке ж, як і у навікули та кратікули, але краї досить порізані і дрібнолопатеві. Види роду мешкають переважно у бентосі морів та континентальних водойм. Особливо високе значення цей рід відіграє у донних угрупуваннях гіпергалійних водойм. Наприклад, у Сакському лікувальному озері у Криму, скупчення клітин Gyrosigma утворюють на дні над лікувальним мулом суцільний щільний шар до кількох см завтовшки.
Стулки видів роду Pinnularia мають форму, наближену до видовжено- еліпсоїдної. Шов у піннулярії дещо хвилястий. З внутрішнього боку стулки панцир має систему грубих потовщень - ребер, які дещо нагадують поверхню пральної дошки, і в оптичний мікроскоп можуть бути помилково прийняті за штрихи. Між ребрами панцир перфорований рядами дрібних ареол. При вигляді з пояска панцир правильно-прямокутний, при чому в цій позиції добре помітно, що ребра є саме системою внутрішніх потовщень стулки, а не її перфорацій. Хлоропластів два, стрічковидні, розташовуються вздовж поясків з протилежних боків стулки і мають дуже порізані краї. У деяких видів протилежні стрічки хлоропласту під гіповальвою з'єднуються невеликим містком, завдяки чому хлоропласт набуває Н-подібної форми. В центральній частині кожної стрічки у багатьох видів розташовується по одному голому еліпсоїдному піреноїду. Оскільки панцири у видів цього роду переважно великі, клітини не здатні утримуватись у товщі води, через що мешкають або у бентосних, або у перифітонних угрупуваннях прісних водойм.
До Bacillariales належать водорості, на стулках яких розташовується по одному з відносно прямому каналовидному шву. Шов бациляріальних складається з вузької зовнішньої щілини, яка сполучається з каналом, розміщеним у внутрішньому потовщенні стулки - кілі. Канал відкривається у порожнину панциру великими отворами - фібулами або кільовими точками. Канал з фібулами є гомологом внутрішньої щілини шва щілиношовних діатомей. За наявністю фібул представники бациляріальних розпізнаються досить легко. Водорості з Bacillariales поширені у водоймах всіх типів та у багатьох ґрунтах. До характерних ґрунтових водоростей належить Hantzschia, водних - Nitzschia.
Стулки у Hantzschia дорзовентральні. Шви обох стулок розташовуються на вентральному боці клітини, поблизу загину стулки. Хлоропластів зазвичай два, лопатевих, розташовуються по різні боки від поперечної площини клітини, і при вигляді зі стулки нагадують дві літери "Н".
У Nitzschia стулки або бісиметричні, або S-подібні. Шви також зміщені до краю стулки, але шов епітеки розташовується на пролежному боці від гіпотеки. Зазвичай види цього роду мають дві пластинчасті цілісні або злегка розсічені пластиди, розміщені по різні боки від поперечної площини клітини. За видовим багатством рід Nitzschia є одним з найчисленніших у відділі. Види з вузькими та дрібними панцирями поширені переважно у планктоні, а з великими та грубими - у бентосі та перифітоні континентальних водойм та морів.
Rhopalodiales належить до групи каналошовних діатомей, у яких кожна стулка має один шов без помітних в оптичний мікроскоп фібул. Клітини ропалодіальних переважно дорзовентральні, місяцеподібно вигнуті, і прикріплюються до субстрату не стулкою, а вентральним боком пояскової зони. Завдяки розширенню пояскових обідків на дорзальному боці (інколи за рахунок вставних обідків), дорзальна пояскова зона є значно ширшою, за вентральну.
Шов починається на полюсах вентрального боку стулки. Далі у одних родів (Epithemia) проходить вздовж вентрального боку і загинається на дорзальний бік; у інших проходить вздовж дорзального боку (Rhopalodia) або майже посередині стулки. У центрі стулки гілки зовнішньої щілини шва утворюють майже непомітний в оптичний мікроскоп центральний вузлик.
Стулки ропалодіальних водоростей з внутрішнього боку мають грубі ребра, між якими розташовується від двох до кількох штрихів, утворених ареолами.
Характерними представниками порядку є перифітонні роди Epithemia та Rhopalodia.
Представників Surirellales інколи називають чотиришовними, оскільки кожна стулка має два каналовидних шва. Вони розташовані на її протилежних боках у зовнішніх складках - крилах. Вважають, що утворення додаткового шва є наслідком поступової зміни полярності панциру внаслідок скорочення довжини стулки і збільшення її ширини з одночасним видовженням полярних щілин шва. Стулки суриреляльних мають складні системи ребер, що підсилюють крила. Між ребрами на стулці знаходяться системи ареол.
Оскільки панцирі представників порядку великі та важкі, ці водорості населяють переважно бентосні угрупування.
Типовий рід порядку - Surirella - має еліпсоїдні або яйцеподібні стулки з добре розвиненими крилами, в яких знаходяться каналовидні шви. На поверхні стулки розташовуються грубі поперечні ребра, що закінчуються на крилах. Клітина має два великих пластинчастих хлоропласти, які розташовуються на периферії клітини таким чином, що один з них притиснутий до епітеки, другий - до гіпотеки. Види роду населяють як моря, так і прісні водойми.
У роду Cymatopleura панцир при вигляді зі стулки перетягнутий і дещо нагадує гітару. Поверхня стулок хвиляста, а крила досить вузькі. Хлоропласт у цієї водорості один, складається з двох пластин, що притиснуті до епі- та гіпотеки, відповідно, і на одному з полюсів з'єднані між собою тонким містком. Види роду населяють прісні водойми, переважно з лужною реакцією середовища.
Поширення та екологічні особливості
До загальних екофізіологічних особливостей Bacillariophyta належать, в першу чергу, їх холодолюбність. Температурний оптимум для більшості видів діатомових водоростей є нижчим від 15° С. Тому діатомові водорості масово вегетують в холодних морях, розвиваються під льодом та на поверхні снігу, часто масово розвиваються взимку. В той же час, незважаючи на холодолюбність, багато видів діатомей є витривалими до дії досить високих температур, через що не припиняють вегетації влітку, і навіть виявляються в гарячих джерелах. Загалом температурний діапазон активної вегетації Bacillariophyta знаходиться в межах від -4° до +50° С при оптимумах різних видів від 0° до 15° С.
У природі діатомові опанували майже всі типи місцезростань, як водних, так і наземних. У крейдяний період (130-100 млн років тому) водорості з Coscinodiscophyceae стали домінуючою групою морського планктону, і наприкінці цього періоду замістили вимираючих на той час коколітофорид (Haptophyta, Coccolithales). Біля 70 млн років тому діатомові почали успішно опановувати прісні водойми. В неогені вони стають одним з провідних відділів континентальних водойм, причому у прісноводному планктоні, на відміну від морського, домінують представники з Fragillariophyceae. Далі, після "винаходу" шва та появою здатності до активного ковзаючого руху, були опновані прісноводні бентосні та перифітонні угрупування, де більшість домінантів належить до Bacillariophyceae. Найпізніше (ймовірно, у плейстоцені) діатомові водорості почали опановувати суходол. Різноманітність суто наземних діатомей невелика, і майже всі наземні види належать до Naviculales та Bacillariales.
Морські діатомеї населяють три основні типи біотопів: планктон, бентос та перифітон. Морський планктон, що розвивається у відкритих акваторіях, далеко від берегів (т.з. пелагічний) представлений переважно видами з Coscinodiscophyceae, які мають тендітні тонкі панцири, що дозволяють парити у водній товщі, але, потрапляючи у прибережну зону, легко руйнуються під дією прибою. У прибережному (неритичному) планктоні розвиваються види з міцнішими панцирами. Зокрема, у Чорному та Азовському морях до домінантів у цьому типі біотопів належать, в першу чергу, види роду Thalassiosira. Крім того, у неритичному планктоні виявляється чимало бентосних та перифітонних діатомей, ресуспендованих у водну товщу прибережними хвилями.
В морському бентосі та перифітоні численними є шовні діатомеї з родів Achnanthes, Cocconeis, Surirella, а також безшовна водорость Licmophora.
Прісноводні діатомеї планктону представлені переважно родами з Fragilariales (Fragilaria, Asterionella, Diatoma та ін.) та видами Nitzschia з тонким видовженими панцирами. В обростаннях найчисленнішими є різноманітні щілиношовні (Cocconeis, Achnathidium, Gomphonema, Rhoicosphenia, Cymbella, Navicula), а також ропалодіальні водорості (Epithemia, Rhopalodia). В бентосі переважають каналошовні з Surirellales та щілиношовнії з товстими, грубими панцирами, зокрема Pinnularia. Деякі з діатомей (наприклад, Melosira) починають свій розвиток у перифітоні, проте далі відриваються від субстрату і переходять до планктонного способу існування.
Значення у природі та житті людини
У біосфері діатомові водорості належать до однієї з провідних груп продуцентів, і відіграють планетарну роль не лише в кругообігу вуглецю та кисню, але й кремнію. Навіть у морях з найбіднішим фітопланктоном біомаса діатомей становить 2-5 тис. т на 1 км акваторії. У планетарному масштабі органічна речовина діатомових водоростей складає біля 50% від всієї біомаси морів та океанів, і їх річна продукція - біля 25% від глобальної планетарної. Щорічно діатомові поглинають зі Світового океану біля 3 млрд. т кремнію.
Панцирі викопних діатомових водоростей утворили осадові породи, відомі під назвою діатоміти, або "інфузорна земля". Діатоміти являють собою легку та пористу, але досить міцну породу, завдяки чому вони використовуються як будівельний матеріал, з них виготовляють тонкі фільтри та абразиви. Завдяки високій адсорбційній здатності їх широко застосовують у хімічній, медичній та харчовій промисловості. З діатомітів, просякнутих нітрогліцерином, виготовляють вибухівку - широко відомий динаміт, винайдений А. Нобелем.
Викопні діатомові водорості використовуються в геології для визначення віку осадових пород (т.з. діатомовий аналіз). У прісних водоймах діатомеї обумовлюють процеси накопичення діатомового сапропелю, який використовується як лікувальний мул.
На основі діатомових водоростей проводиться оцінка якості води за т.з. діатомовими індексами. Ця методика стандартизована в країнах Європейської спільноти, і є обов'язковою при проведенні будь-яких екологічних експертиз річок та озер на території Західної Європи.
Відомі випадки, коли діатомові, розвиваючись масово, викликають "цвітіння води". Особливо часто збудниками "цвітіння" у прісних водоймах стають центричні діатомеї, зокрема види роду Cyclotella. В цих випадках вода набуває брудно-жовтого кольору та неприємного запаху риб'ячого жиру. В останні роки з'явились дані, що в деяких випадках "цвітіння води", обумовлене розвитком діатомей, може бути помірно токсичним.
Місце Bacillariophyta у системі органічного світу
Хоча належність діатомових водоростей до філи тубулокристат-страменопілів сумнівів не викликає, система родинних зв'язків цього відділу з іншими хромофітовими водоростями не є досить визначеною. Найбільш ймовірними близькими родичами діатомей вважають золотисті або жовтозелені водорості, у яких чітко окреслюється тенденція просякування оболонок кремнеземом, утворення кремнеземових лусочок або цист зі зкрем'янілими покривами.
1 За оцінками Ф. Раунда та Р. Крауфорда (1990), які приймають монотипічну концепцію виду у діатомей, у відділі відомо біля 100 тис. видів. За підрахунками Т. Нортона зі співавторами (1996), очікуване видове багатство цього відділу знаходиться у діапазоні від 100 тис. до 10 млн. видів.