Альгологія - Костіков І.Ю. - 2009-2013
Розділ 18. Диктіохофітові водорості – Dictyochophyta
Диктіохофітові водорості - це специфічна група хроміст, яку донедавна включали до відділу Chrysophyta, розглядаючи їх як окремий самостійний порядок - Dictyochales з класу примнезіофіцієвих та кілька окремих родів в системі класу хризофіцієвих. У самостійний таксон хроміст, ранг якого відповідає щонайменш класу, а вірогідно - відділу, диктіохофіти були виділені внаслідок інтенсивних електронно-мікроскопічних досліджень 80-90-х років. Вже перші молекулярно-філогенетичні дослідження гену, що кодує 18S РНК диктіохофітів підтвердили обгрунтованість такого підходу і навіть більше - показали, що диктіохофіти є такою ж незалежною філою в межах хроміст, як і діатомові або рафідофітові водорості, тобто з філогенетичних позицій їх ранг відповідає рангу відділу. В межах диктіохофітів чітко окреслені два класи - диктіохофіцієві (силікофлагеляти) та пединелофіцієві водорості (рис. 18.1).

Рис. 18.1. Диктіохофітові водорості двох головних еволюційних ліній відділу: 1 - Dictyocha, 2 - Pedinella (за Матвієнко, 1965).
Сучасні диктіохофіти об’єднують біля 40 видів мікроскопічних одноклітинних водоростей з монадною або амебоїдною будовою. Відома також значна кількість викопних диктіохофітів; найдавніші представники мають вік біля 120 млн. років. Поширені диктіохофіти переважно у планктоні морів та прісних водойм.
Систематичні ознаки відділу
Пігменти та запасні поживні речовини
Біохімічними ознаками відділу є наявність хлорофілів "а" та "c", які маскуються додатковими пігментами, в першу чергу - фукоксантином. Через це хлоропласти диктіохофітів забарвлені у жовтий або світло-коричневий колір. З інших ксантофілів виявлено діатоксантин, діадіноксантин, лютеін. З каротинів знайдено лише β-каротин. Запасними поживними речовинами є олія та хризоламінарин.
Цитологічні ознаки
Клітинні покриви представлені двома основними варіантами: у силікофлагелят - це плазмалема, над якою знаходиться кремнеземовий скелет, одягнений слизовою капсулою. У пединелофіцієвих водоростей клітинні покриви представлені плазмалемою, звичайно вкритою субмікроскопічними лусками.
Ядерний апарат. Диктіохофітові водорості одноядерні. Ядро еукаріотичного типу, відмежовано від цитоплазми двомембранною оболонкою, з одним або кількома ядерцями. З апікального боку до ядерної оболонки прикріплюються два базальних тіла джгутиків. До ядерної оболонки також безпосередньо прилягає комплекс Гольджі; в цьому місці зовнішня ядерна мембрана здатна відщеплювати пухирці, які надалі вбудовуються у цистерни комплексу Гольджі. Крім того, у диктіохофітів наявні особливі цитоплазматичні мікротрубочкові структури, які відходять від поверхні ядра і утвоюють у цитоплазмі додаткові опірні елементи, а у амебоїдних форм заходять у ризоподії і приймають участь у їх русі.
Мітохондрії. На ультратонких зрізах мітохондріальні профілі мають трубчасті кристи.
Фотосинтетичний апарат. Переважна більшість диктіохофітів є фотоавтотрофами або міксотрофами, хоча серед пединелових водоростей відомі чотири облігатно гетеротрофних роди. У здатних до фотосинтезу представників клітина звичайно містить кілька пристінних хлоропластів. Кожний хлоропласт вкритий чотирьохмембранною оболонкою. Дві внутрішні мембрани є власними мембранами хлоропласту, дві зовнішні мембрани утворюють хлоропластний ендоплазматичний ретикулюм. Між власними хлоропластними мембранами та мембранами хлоропластного ендоплазматичного ретикулюму є досить великий перипластидний простір з перипластидним ретикулюмом та 80S рибосомами.
Під хлоропластною оболонкою розташовується оперезуючий тилакоїд, далі - звичайні тилакоїди, зібрані переважно по три. В стромі хлоропласту також міститься 70S рибосоми, хлоропластна ДНК (рис. 18.2).
Джгутиковий апарат, з одного боку, має типові риси, притаманні хромістам, з іншого боку, ряд специфічних ознак, що яскраво відокремлюють диктіохофітів від інших відділів хроміст. Так, типовими ознаками є дводжгутиковий гетероконтний, гетероморфний та гетеродинамічний план будови. Звичайно короткий джгутик повністю редукований і від нього зберігається лише базальне тіло. Вийнятком є лише зооспори диктіохофіцієвих водоростей. Довгий джгутик несе два ряди типових для хроміст тричастних мастигонем (рис. 18.2).
Унікальними особливостями джгутикового апарату диктіохофітів є повна відсутність мікротрубочкових джгутикових коренів та прикріплення базальних тіл джгутиків безпосередньо до ядерної мембрани. Ознакою, що зближує джгутиковий апарат диктіохофітів з золотистими, жовтозеленими та еустигматофітовими водоростями є наявність проміжної спіралеподібної структури, що розміщується у перехідній зоні джгутика - між аксонемою та базальним тілом.
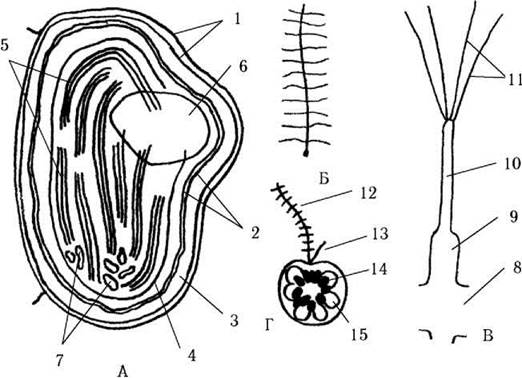
Рис. 18.2. Схема будови хлоропласту (А), джгутику (Б), мастигонеми (В) та зооспори (Г) диктіохофіцієвих водоростей (на прикладі Dictyocha): 1 - зовнішні мембрани хлоропласту; 2 - внутрішні мембрани хлоропласту; 3 - перипластидний простір; 4 - оперезуючий тилакоїд; 5 - ламели, що складаються з зібраних по три тилакоїдів; 6 - напівзанурений піреноїд; 7 - хлоропластна ДНК, що складається з кількох замкнених у кільце ізольованих сегментів; 8 - аксонема джгутика; 9 -11 - мастигонема (9 - розширена частина, 10 - проміжна ділянка, 11 - термінальні волоски); 12 - пірчастий джгутик зооспори; 13 - гладенький джгутик; 14 - хлоропласти; 15 - вакуоля (за Moestrup, Thompson, 1990; Van Valkenburg, 1971).
Розмноження
Розмноження відбувається або вегетативним поділом клітин надвоє, або за допомогою зооспор. У пединелових водоростей відомі також цисти, які зовні нагадують цисти золотистих водоростей, проте не просякнуті кремнеземом.
Система відділу
Відділ включає два класи, поділ на які відбувається за особливостями будови клітин - Dictyochophyceae (силікофлагеляти) та Pedinellophyceae. Основні відміни між цими класами полягають у загальному плані організації клітини, будові покривів, кількості та роміщені комплексу Гольджі, деталях організації фотосинтетичного апарату, способі розмноження та наявності цист.
Клас Dictyochophyceae
План організації клітини. Клітини диктіохофіцієвих представлені одноджгутиковими монадами. Протопласт клітини диференційований на дві зони - центральну перинуклеарну та периферичну. Периферична зона складається з числених радіально розміщених тяжів цитоплазми, які анастомозують між собою та розширюються на верхівці. В цих розширеннях розташовуються хлоропласти та мітохондрії (рис. 18.3). У перинуклеарній зоні міститься ядро, комплекси Гольджі, від центральної зони відходять джгутик, інколи - псевдоподія, яку називають щупальцем.
Клітинні покриви утворює слизова капсула, всередині якої знаходиться протопласт клітини, відмежований від слизу лише плазмалемою. У периферичній частині слизової капсули, проте назовні від плазмалеми, розміщується кремнеземовий скелет. Скелет має вигляд кошика, який утворений з'єднаними між собою полими кремнеземовими трубкам (рис. 18.4).

Рис. 18.3. Будова клітини диктіохофіцієвих водоростей (на прикладі роду Dictyocha, вид зверху): 1 - межа слизової капсули; 2 - аморфний слиз; 3 - псевдоподія; 4 - цитоплазматичний тяж, що з’єднує периферичну та перинуклеарну зони клітини; 5 - периферична зона протопласту; 6 - елемент кремнеземового скелету; 7 - вакуоля; 8 - пери- нуклеарна зона протопласту; 9 - джгутик; 10 - ядро; 11 - ядерце; 12 - комплекс Гольджі; 13 - хлоропласт; 14 - мітохондрія; 15 - піреноїд (за Van Valkenburg, 1971).

Рис. 18.4. Кремнеземовий скелет Dictyocha, вид збоку (за Deflandre, 1952).
Ядро містить одне велике ядерце; навколо ядерної оболонки розташовується кілька (іноді - до кількох десятків) комплексів Гольджі.
Фотосинтетичний апарат силікофлагелят, у порівнянні з пединелофіцієвими, має три особливості: по-перше, хлоропласти числені (до кількох десятків), по-друге, хлоропластна оболонка не переходить у ядерну оболонку, по-третє, хлоропластна ДНК розсіяна по стромі у вигляді кількох фрагментів.
Джгутикові стадії представлені вегетативними клітинами та зооспорами.
Розмноження у відомих представників класу відбувається за допомогою зооспор. Зооспори диктиохофіцієвих водоростей голі, позбавлені кремнеземового скелету, і, на відміну від вегетативних клітин, несуть два гетероконтні та гетеродинамічні джгутики, що виходять з апікальної заглибини (див. рис. 18.2). Кожна зооспора проростає у звичайну вегетативну клітину. Цисти у диктиохофіцієвих не виявлені.
Система класу. В межах класу представлено один порядок - Dictyochales та одну родину - Dictyochaceae. У сучасній флорі відомо два роди - Dictyocha та Distephanus (рис. 18.5).
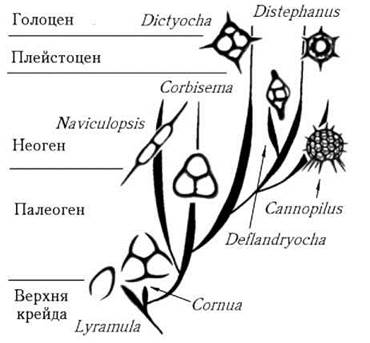
Рис. 18.5. Схема еволюційного розвитку основних родів диктіохофіцієвих водоростей (за Глезер, 1977).
Крім того, до диктіохофіцієвих за деякими ультраструктурними ознаками та ступенем подібності гену, що кодує 18S РНК є близькими морські пікопланктонні водорості Pelagococcus та Pelagomonas, які деякі автори розглядають або як окремий порядок диктіохофіцієвих, або як окремий клас диктіохофітів, або навіть як окремий відділ хроміст, споріднений з Dictyochophyceae.
Поширення, екологічні особливості та значення. Диктиохофіцієві водорості, незважаючи на їх низьку різноманітність, надзвичайно широко розповсюджені у морях та океанах високих широт. Чисельність сучасних силікофлагелят у планктоні звичайно невисока, і поступається чисельності діатомових, динофітових та примнезіофітових водоростей. Проте наприкінці 80- років з’явилися тривожні дані про нове токсичне "цвітіння", збудником якого стала силікофлагелята Distephanus speculum.
Однією з яскравих екологічних особливостей диктіохофіцієвий є їх виражена психрофільність (холодолюбність). Температурний оптимум вегетації для представників Dictyochophyceaeзнаходиться у діапазоні 5-12 °С. За цією рисою силікофлагеляти близькі до діатомових водоростей.
Найбільш давні викопні рештки диктиохофіцієвих водоростей датуються віком близько 120 млн років, що може вказувати на відносну "молодість" цих організмів. Вважають, що еволюція силікофлагелят була пов’язана з поступовим ускладненням кремнеземового скелету (рис. 18.5). Оскільки скелет силікофлагелят добре зберігається у викопному стані, ці водорості використовуються в геологічній практиці - для визначення віку осадових порід та реконструкції температурних умов Світового океану в найближчі до нас геологічні епохи.
Клас Pedinellophyceae
План організації клітини. Клітини пединелофіцієвих водоростей монадні, одноджгутикові. Джгутик виходить із апікальної заглибини. Звичайно на передньому кінці клітина здатна утворювати псевдоподії, а на задньому - стебельце, яке у частини видів здатне скорочуватись. Протопласт не диференційований на периферичну та перинуклеарну зони (рис. 18.6).
Клітинні покриви представлені плазмалемою, на поверхні якої відкладаються субмікроскопічні лусочки. Лусочки утворюються комплексом Гольджі і виштовхуються назовні на задньому кінці.
Ядро безпосередньо пов’язано з зовнішньою мембраною хлоропласта, базальними тілами джгутиків та тритубулярними цитоскелетними структурами. До базальної сторони ядра прилягає один комплекс Гольджі. Зовнішня мембрана ядра з цього боку здатна відокремлювати пухирці, які надалі вбудовуються у комплекс Гольджі.

Рис. 18.6. Будова клітини та цисти пединелофіцієвих водоростей (на прикладі Mesopedinella arctica): 1 - лусочки, 2 - джгутикова субапікальна заглибина, 3 - базальне тіло довгого джгутика, 4 - базальне тіло редукованого короткого джгутика, 5 - проміжна спіралеподібна структура, 6 - тричасна мастигонема, 7 - параксилярний тяж, 8 - аксонема джгутика, 9 - тритубулярна цитоскелетна структура, 10 - мітохондріальний профіль з трубчастими кристами, 11 - ядро, зовнішня мембрана якого повязана з зовнішньою мембраною хлоропласту, 12 - хлоропласт, 13 - ядерно-мембранні пухирці, 14 - цистерни комплексу Гольджі, 15 - вакуоль, 16 - пухирець комплексу Гольджі з платівкою клітинного покриву, 17 - зона утворення стебельця та виходу назовні платівок клітинних покривів (за Daugbjerg, 1996).
Фотосинтетичний апарат у деяких представників редукований. У фотоавтотрофних пединелофіцієвих він, найчастіше включає шість парієтальних хлоропластів. Зовнішня мембрана хлоропласту, принаймі у пединеляльних водоростей, безпосередньо переходить у зовнішню ядерну мембрану. Хлоропластна ДНК не розсіяна по стромі, а зконцентрована у вигляді однієї замкненої у кільце молекули.
Монадні клітини завжди одноджгутикові. Основу цитоскелету складає опірна мікротубулярна система з тримікротрубочкових структур. Тримікротрубочкові структури утворюють систему ніжок та кільця. Один кінець ніжки прикріплений до ядерної мембрани, протилежний кінець з'єднується з шістьма-дванадцятьма триплетами мікротрубочок, що замикаються у кільце навколо субапікальної заглибини.
Розмноження відбувається поділом клітин надвоє у рухливому стані. У видів, що мають стебельце, водночас з поділом клітини спостерігається його продольне розщеплення. Несприятливі умови пединелофіцієві переносять у стані цист (рис. 18.6). Цисти більш-менш сферичні, не просякуті кремнеземом, мають отвір, закритий пробкою та одягнені тришаровою оболонкою, яка оточена шаром лусочок.
Система класу. В межах класу представлено два порядки - Pedinellales та Rhizochromulinales. Порядки розділяються, в першу чергу, за ультраструктурними особливостями. Так, у Rhizochromulinales наявні деякі риси схожості з диктіохофіцієвими водоростями: джгутик не має параксилярного тяжа, клітина не вкрита субмікроскопічними лусками, зовнішня хлоропластна мембрана не продовжується у ядерну, відсутнє стебельце. За іншими ознаками цей порядок відповідає ознакам класа Pedinellophyceae (включаючи відсутність кремнеземового скелету). Молекулярно-філогенетичні дослідження 18S РНК підтвердили філогенетичну спорідненість Pedinellales та Rhizochromulinales між собою. Два представника класу - Pedinella та Mesopedinellaпредставлені на рис. 18.1 та 18.6.
Поширення та екологічні особливості. Пединелофіцієві водорості поширені переважно у холодних морях, рідше зустрічаються у прісних водоймах. Один вид - Pedinella hexocostata - знайдений в Україні в озерах Донеччини.
Таким чином, відділ Dictyochophyta демонструє унікальний тип клітинної організації, що відрізняє його від інших відділів хроміст - в першу чергу, це відсутність джгутикових коренів, безпосередній зв'язок базальних тіл з ядерною оболонкою та специфічний тип цитоскелету. Ультрастуктурні дані про самобутність та філогенетичну самостійність диктіохофітів добре узгоджуються з результатами молекулярно-філогенетичних реконструкцій родинних зв'язків хроміст.