Альгологія - Костіков І.Ю. - 2009-2013
Розділ 20. Гаптофітові водорості – Haptophyta
Гаптофітові водорості можна вважати "живими свідками" тих часів, коли почалась дивергенція тубулокристат і платикристат. Сьогодні відділ нараховує біля 500 видів з 80 родів. Переважна більшість гаптофітових - це морські одноклітинні організми.
Систематичні ознаки відділу
Пігменти та запасні поживні речовини
Хлоропласти примнезіофітів звичайно забарвлені у жовтий або коричневий колір. Забарвлення визначається наявністю хлорофілів а та с, які маскуються жовтими ксантофілами: фукоксантином та його дериватами - гексанолокси- або бутанолоксифукоксантином, та діадіноксантином й діатоксантином. З каротинів виявлено лише Р-каротин.
Основним продуктом асиміляції є хризоламінарин, що відкладається поза межами пластид. У гаптофітових з порядку Pavlovales виявлено також парамілон, який раніш вважали специфічним продуктом асиміляції лише евгленофітових водоростей.
Цитологічні ознаки
З точки зору цитології відділ цікавий тим, що демонструє велику кількість модифікацій ендоплазматичного ретикулюму, пов'язаних майже з усіма основними органелами - ядром, хлоропластом, покривами, комплексом Гольджі, особливою структурою - гаптонемою, частково - із джгутиковим апаратом, а також з процесом мітозу.
Клітинні покриви. Клітини вкриті плазмалемою, на поверхні якої знаходиться один чи кілька шарів субмікроскопічних целюлозних лусочок або гранул, а з боку цитоплазми розташовується одна велика оперезуюча цистерна ендоплазматичної сітки, що утворює два додаткові внутрішні шари покривних мембран (рис. 20.1). Лусочки та гранули прикріплюються одна до одної та до плазмалеми за допомогою кислих полісахаридів. У деяких видів лусочки просякуються вапном, утворюючи органомінеральні покривні елементи, що отримали назву коколітів. Морфологія лусочок використовується як важлива таксономічна ознака переважно на видовому рівні (рис. 20.2).
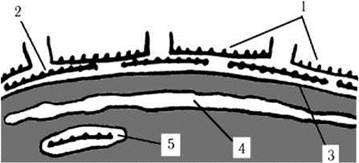
Рис. 20.1. Схема будови клітинних покривів гаптофітових: 1, 2 - поверхневі лусочки, 3 - плазмалема, 4 - оперезуюча цистерна ендоплазматичної сітки, 5 - пухирець комплексу Гольджі, що транспортує до поверхні нову лусочку.

Рис. 20.2. Лусочки різних видів роду Chrysochromulina (за Esperland, Throndsen, 1986; Birkhead, Pienaar, 1995; Eikrem, 1996).
Ядерний апарат. Ядро має еукаріотичну будову, проте з багатьма специфічними особливостями. Так, зовнішня ядерна мембрана безпосередньо переходить у зовнішню мембрану хлоропласту (рис. 20.3). Крім того, вона утворює інвагінації, на яких розміщуються цитоплазматичні рибосоми. Звичайно у гаптофітових не спостерігається чітко окреслене ядерце. В метафазі не розрізняються окремі хромосоми, а весь хроматин конденсується в одну щільну хроматинову платівку. Кінетохори не виявлені. Відсутні також і центриолі, а функцію центру організації мікротрубочок веретена поділу виконує система пухирців комплексу Гольджі, або (у водоростей з порядку Pavlovales) електронно-щільні структури, розташовані на кінці мікрофібрилярного кореню довшого джгутика.
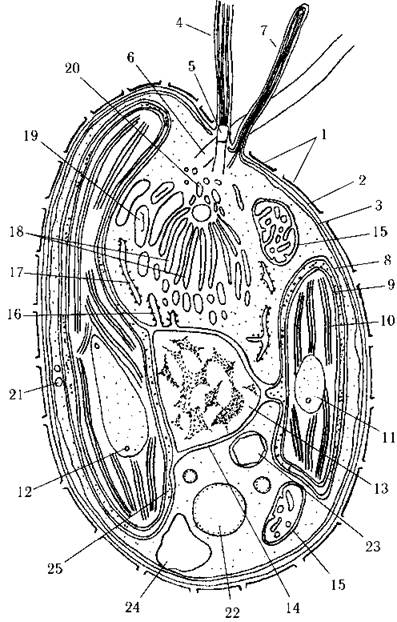
Рис. 20.3. Схема будови клітини гаптофітових водоростей: 1 - поверхневі лусочки, 2 - плазмалема, 3 - периферична оперезуюча цистерна ендоплазматичного ретикулюму, 4 - джгутик, 5 - перехідна зона джгутику, 6 - базальне тіло, 7 - гаптонема, 8 - зовнішня мембрана оболонки хлоропласту (хлоропластний ендоплазматичний ретикулюм), 9 - внутрішні мембрани оболонки хлоропласту, 10 - тритилакоїдна ламела, 11 - піреноїд, 12 - тилакоїд, що пронизує піреноїд, 13 - ядро, 14 - зовнішня мембрана оболонки ядра, що продовжується у зовнішню мембрану хлоропласту, 15 - мітохондрія з трубчастими кристами, 16 - інвагінації зовнішньої мембрани ядерної оболонки (ядерний ендоплазматичний ретикулюм), 17 - канал ендоплазматичного ретикулюму, 18 - цистерни комплексу Гольджі, 19 - пухирець комплексу Гольджі з поверхневою лусочкою, 20 - звичайні пухирці комплексу Гольджі, 21 - слизові тільця, 22 - хризоламінарин, 23 - поліфосфатне тіло, 24 - вакуоль, 25 - перипластидний простір (узагальнено за мікрофотографіями Green, Hibberd, Pienaar, 1982; Birkhead, Pienaar, 1995, Eikrem, 1996).
Гаптофітовим водоростям властивий відкритий, напівзакритий або закритий мітоз. Його особливості полягають у тому, що у метафазі хромосоми конденсуються у єдину масивну платівку, пронизану каналами. По каналах проходять пучки мікротрубочок веретена поділу. До веретена прилягають два комплекси Гольджі, які повністю відокремлюють веретено від базальних тіл джгутиків. У телофазі з цистерн комплексу Гольджі та полярних пухирців формується нова ядерна оболонка, а цитокінез відбувається за рахунок кільцевої борозни, яку утворює плазмалема.
За аналізами подібності нуклеотидних послідовностей цілого ряду ядерних генів показано, що ядерний геном гаптофітових, з одного боку, близький до ядерних геномів криптофітових, зелених, червоних та глаукоцистофітових водоростей, з іншого боку має певні риси схожості з геномами плазмодіофороміцетових слизовиків та сонячників-центрохелідій.
Фотосинтетичний апарат. Хлоропласти гаптофітових одягнені чотирма мембранами, з яких дві зовнішні складають хлоропластну ендоплазматичну сітку, а дві внутрішні - власне хлоропластну оболонку. Зовнішня мембрана безпосередньо продовжується у зовнішню мембрану ядра. Між внутрішніми та зовнішніми мембранами знаходиться перипластидний простір з власними 80Sрибосомами.
Тилакоїди у стромі хлоропласту зібрані по три, оперезуючої ламели немає. Досить часто наявний голий піреноїд. Хлоропластна ДНК складається з численних фрагментів, дифузно розсіяних по сторомі хлоропласту.
Вічко є тільки у водоростей з порядку Pavlovales. Воно розташовано у хлоропласті, складається з одного шару дрібних глобул, що безпосередньо прилягають до хлоропластної оболонки, і не зкоординовано із джгутиками.
Таким чином, цитологічні ознаки вказують на вторинно-симбіотичне походження пластид гаптофітових. За молекулярно-філогенетичними реконструкціями, пластиди примнезіофітів виводяться від червоних водоростей.
Мітохондрії гаптофітових мають трубчасті кристи, подібні до крист водоростей-хроміст та інших страменопілів. Дослідження гену, що кодує першу субодиницю мітохондріального цитохрому с підтвердили припущення про спорідненість мітохондрій Haptophyta з мітохондріями тубулокристат, зокрема - еустигматофітових водоростей.
Надзвичайно цікавим виявився також факт наявності у мітохондріальної ДНК гаптофітових деривату генетичного коду тваринного типу. Так, якщо в мітохондріях вищих рослин триплет UGAвиконує функцію стоп-кодону при біосинтезі білку, то в мітохондріях багатоклітинних тварин, грибів та червоних водоростей цей триплет кодує амінокислоту триптофан. У гаптофітових UGAтакож кодує триптофан, і не є стоп-кодоном.
Джгутиковий апарат. Монадні стадії представлені вегетативними клітинами, зооспорами та гаметами. Рухливі клітини мають два джгутики рівної або нерівної довжини, що виходять із субапікальної або латеральної заглибини. Звичайно джгутики гладенькі, проте у водоростей з порядку Pavlovales обидва джгутики мають мастигонеми простої (не страменопільної) будови, причому довший джгутик вкритий також субмікроскопічними гранулами.
На відміну від переважної більшості інших відділів водоростей, перехідна зона джгутика гаптофітових водоростей не має універсальної будови. У Haptophyta відомо три основні структури перехідної зони: а) аксосома — велика капелюхоподібна втулка; б) спіралеподібна структура; в) система проміжних кілець. У деяких видів водночас наявні всі ці структури (рис. 20.4а), у інших - лише дві з них або тільки якась одна.
Крім джгутиків, у частини гаптофітових водоростей є унікальна джгутикоподібна нерухома органела - гаптонема. Вважають, що гаптонема виконує функції прикріплення клітин до субстрату. Вона являє собою виріст плазмалеми, в якому розташована одна цистерна ендоплазматичної сітки, що обгортає 6-7 мікротрубочок. Останні є продовженням одного з джгутикових коренів (рис. 20.4б).
У Haptophyta, за винятком роду Pavlova, коренева система джгутиків включає три основних мікротрубочкових кореня (вони відходять у цитоплазму від базальних тіл джгутиків), один-два великих додаткових (т.з. кристалоподібних) кореня, які приєднуються до основних, та один гаптонемовий корінь, що утворений мікротрубочками власне гаптонеми. Зв'язок коренів між собою та з базальними тілами здійснюється за допомогою скоротливого білку центрину (рис. 20.4: в,г).

Рис. 20.4. Будова джгутика (а), гаптонеми (б) та кореневої системи джгутикового апарату (в - вигляд збоку, г - вигляд зверху) гаптофітових водоростей: 1 - базальне тіло, 2 - аксонема, 3 - центральний дублет мікротрубочок, 4 - периферичний дублет мікротрубочок, 5 - спіралеподібна структура, 6 - аксосома, 7 - проміжні кільця, 8 - плазмалема, 9 - периферична цистерна ендоплазматичного ретикулюму, 10 - поверхневі лусочки, 11 - цистерна ендоплазматичного ретикулюму у гаптонемі, 12 - мікротрубочки гаптонеми, 13 - цистерна ендоплазматичного ретикулюму, що при основі гаптонеми зміщена на один бік, 14, 15 - базальні тіла джгутиків (14 - лівого, 15 - правого), 16 - 18 - основні мікротубулярні корені (16 - перший, 17 - другий, 18 - третій), 19 - гаптонемовий корінь, 20 - додатковий корінь другого мікротубулярного кореню, 21, 22 - мікрофібрилярні корені першого та третього основних коренів, 23 - система фібрилярних з'єднань базальних тіл (схематизовано за Beech, Wetherbee, 1988; Roberts, Mills, 1992; Gregson, Green, Leadbeater, 1993).
Інші органели. Між базальними тілами джгутиків та ядром звичайно розташовується один великий комплекс Гольджі. Він, на відміну від хроміст, не притиснутий до ядерної мембрани. Крім звичайних функцій, комплекс Гольджі приймає участь в утворенні лусочок, що далі транспортуються у пухирцях назовні і відкладаються на поверхні плазмалеми.
На периферії клітини досить часто можна спостерігати слизові тіла, на її задньому кінці - великі краплини хризоламінарину, а у павловальних водоростей - також поліфосфатні тіла та зерна парамілону.
Плани будови тіла
Переважна більшість гаптофітових мають монадну будову. Рідше зустрічаються кокоїдні форми. Гаметоспорфіти деяких примнезіальних, ізохризидальних та коколітальних водоростей мають нитчастий тип структури. За несприятливих умов більшість видів Haptophyta здатні втрачати джгутики та переходити у пальмелеподібний стан.
Розмноження та життєві цикли
Гаптофітові розмножуються переважно вегетативним поділом клітин на дві. Зрідка зустрічається також розмноження за допомогою зооспор. У небагатьох видів описаний ізогамний статевий процес.
У більшості видів життєвий цикл являє собою цикломорфоз. Наприклад, у життєвому циклі морської водорості Phaeocystis відбувається зміна монадної та кокоїдної стадій. При цьому кокоїдні клітини здатні утворювати мікро- або макроскопічні слизові колонії, а при руйнуванні колоній повертатися у монадний стан (рис. 20.5: а).
У еугамних видів життєвий цикл досить складний, він відбувається з гетероморфною зміною поколінь та споричною редукцією, і, таким чином, є гаплодиплофазним.
Наприклад, у роду Pleurochrysis монадні вегетативні диплоїдні клітини являють собою спорофіт, відомий як стадія " Hymenomonas". Вони вкриті коколітами, розмножуються поділом надвоє або за допомогою зооспор. За певних умов відбувається мейоз, внаслідок якого утворюються чотири гаплоїдні зооспори, що проростають у гаплоїдний розгалужений нитчастий гаметоспорфіт, раніш описаний як самостійний рід "Apistonema". Гаметоспорофіт відтворює себе зооспорами або дає ізогамети. Після копуляції зигота без періоду спокою проростає у монадну диплоїдну коколітофориду (рис. 20.5: б). Зооспори, що продукує спорофіт, мають коротку дуже редуковану гаптонему; у зооспор гаметоспорофіту гаптонема може бути наявна або відсутня.

Рис. 20.5. Життєві цикли гаптофітових водоростей. а - цикломорфоз (на прикладі Phaeocystis pouchetii), б - гаплодиплофазний життєвий цикл з гетероморфною зміною поколінь (на прикладі Pleurochrysis carterae) (за Kommann, 1955; Lee, 1980).
Система відділу
За загальним планом будови клітин у відділі простежуються дві чітко окреслені лінії. Перша лінія об’єднує всі типові гаптофітові, друга - представників роду Pavlova. Суттєві відміни між цими лініями полягають у здатності накопичувати парамілон, наявності мастигонем, субмікроскопічних лусочок на джгутиках, вічка, у положенні ядра, деталях будови цитоскелету. У зв’язку з цим висловлювались пропозиції розділити відділ на два класи або підкласи. Проте молекулярно-філогенетичні реконструкції як за ядерним геномом, так і за геномом хлоропласту поки що не підтвердили обґрунтованості такого поділу. Тому сьогодні у межах відділу виділяють лише один клас - Haptophyceae, який поділяють на чотири порядки - Isochrysidales, Coccolithales, Prymnesiales, Pavlovales. В основі поділу на порядки лежить не тільки план будови клітин, але й особливості будови клітинних покривів та наявність гаптонеми (табл. 20.1).
Табл. 20.1. Основні ознаки різних порядків Haptophyceae
гаптонема |
коколіти |
стигма, мастигонеми, субмікроскопічні лусочки на джгутику, парамілон |
|
Prymnesiales |
є |
відсутні |
відсутні |
Isochrysidales |
відсутня |
відсутні |
відсутні |
Coccolithales |
відсутня |
є |
відсутні |
Pavlovales |
є |
є |
є |
1. Prymnesiales. Об’єднує одноклітинні та колоніальні монадні та амебоїдні рівноджгутикові водорості з довгою або короткою гаптонемою. Коколіти відсутні, на поверхні плазмалеми є лише мікроскопічні целюлозні лусочки. Представники порядку мешкають як у морському планктоні, так і у прісних континентальних водоймах. Деякі примнезіальні водорості здатні викликати токсичне "цвітіння" води. Найбільш поширеними є монадні водорості, в першу чергу - види родів Chrysochromulina, Prymnesium, Corymbellus, Phaeocystis. Прикладом амебоїдних примнезіальних є рід Platychrysis (рис. 20.5; 20.6).
2. Isochrysidales. Об'єднує одноклітинні та колоніальні монадні та кокоїдні водорості, що не мають гаптонеми і не утворюють коколітів. Монадні клітини ізохризідальних мають однакові або злегка нерівні джгутики. Мешкають переважно в морях. Характерними родом є Chrysotila (рис. 20.6).
3. Coccolithales (Coccosphaerales, Coccolithophorales). Об’єднує т.з. коколітофорид - монадні або кокоїдні водорості, що позбавлені гаптонеми, проте мають кальцифіковані поверхневі лусочки - коколіти. Поширені переважно в морях. Коколітофориди панували наприкінці мезозою, у кайнозої більшість видів вимерла. З сучасних родів типовими монадними представниками є Pleurochrysisта Hymenomonas (рис. 20.5; 20.6), кокоїдний тип структури властивий роду Emiliania (рис. 20.7).
4. Pavlovales. Об'єднує монадні водорості з особливим, "павловальним" планом будови клітин. Специфічними рисами порядку є наявність двох джгутиків нерівної довжини (обидва - з простими мастигонемами, а довший також з субмікроскопічними лусочками), стигми, парамілону, особливих "шишкоподібних" коколітів, редукованої гаптонеми. Ядро займає апікальне положення, а мікрофібрилярний джгутиковий корінь виконує функції центру організації мікротрубочок (рис. 20.7). Поширені у планктоні морів та естуаріїв. Включає поки що лише рід Pavlova.
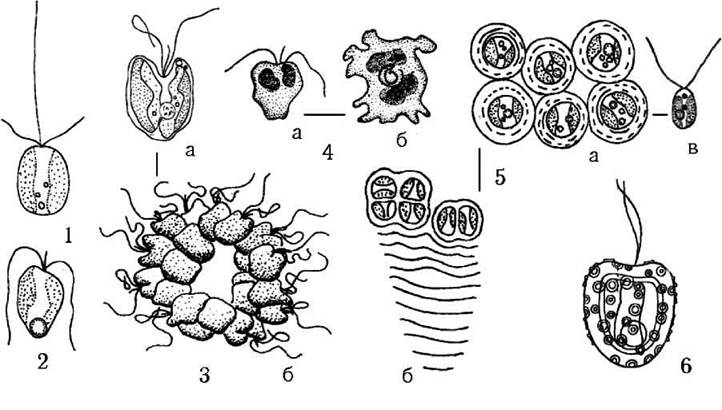
Рис. 20.6. Деякі представники гаптофітових водоростей: 1 - Chrysochromulina sp., 2 - Prymnesium parvum, 3 - Corymbellus aureus (а - окрема клітина, б - колонія), 4 - Platychrysis pigra (а - монадна клітина, б - амебоїдна), 5 - Chrysotila lamellosa (а - поодинокі клітини у пальмелеподібному стані, б - колонія, в - зооспора), 6 - Hymenomonas roseola (за Матвієнко, 1965; Green, 1976; Billard, Gayral, 1972; Топачевский, Масюк, 1984).

Рис. 20.7. Коколітофорида Emiliania huxleyi (ліворуч) та схема будови клітини Pavlova (праворуч) (за Klaveness, 1972, 1976; схематизовано за van der Veer, 1972, 1976).
Поширення, екологія та значення
Гаптофітові водорості мешкають переважно в морях, і є характерним компонентом планктонних угруповань. У товщі води клітини монадних гаптофітових здійснюють добові міграції: удень вони зосереджуються у теплих поверхневих шарах еуфотичної зони, на глибині 30-80 см, вночі спускаються у більш холодні і збагачені біогенними елементами глибинні шари, в окремих випадках сягаючі термоклину (на глибини до 3-5 метрів).
Гаметоспорофіти деяких коколітофорид (зокрема, нитчаста форма "Apistonema" роду Pleurochrysis) розвиваються у бентосі або перифітоні, на поверхні кальцифікованих субстратів - вапняках, крейдяних скелях, черепашках молюсків тощо.
Особливо численні гаптофітові (в першу чергу - з Coccolithales) у тропічних та субтропічних морях, де разом з динофітами є основними продуцентами фітопланктону. Менша кількість Haptophytaзустрічається морях помірних та помірно-холодних поясів. Деякі види, наприклад Emiliania huxleyi, є космополітами.
Гаптофітові можуть спричинювати "цвітіння" води. Так, Emiliania huxley та Phaeocystis pouchetii періодично викликають нетоксичні "цвітіння" у північній Атлантиці вздовж узбережжя Європи. Сьогодні встановлено, що масовий розвиток цих видів впливає на погоду та клімат. Такий вплив пов’язаний із здатністю клітин Emiliania та Phaeocystis в процесі метаболізму виділяти в атмосферу леткий диметилсульфід. Ця речовина після фотохімічної трансформації в верхніх шарах атмосфери утворює в хмарах центри конденсації вологи, що призводить до випадіння кислотних дощів. Інший збудник нетоксичного "цвітіння" - колоніальна водорость Corymbellus aureus - регулярно масово розвивається у проливі Ла-Манш.
Перше токсичне примнезіофітове "цвітіння" у морях зареєстровано наприкінці 80-х років - воно охопило узбережжя Норвегії та Швеції, тривало протягом майже двох місяців і спричинило масову загибель лососевих риб на морських фермах та у рибоводних господарствах. Збудником "цвітіння" стала Chrysochromulina polylepis.
Збудником небезпечного токсичного "цвітіння" солонуватих континентальних водойм та естуаріїв є Prymnesium parvum. При масовому розвитку він виділяє у воду суміш протеофосфоліпідів, що отримала назву іхіотоксину. Іхтіотоксин після активації іонами натрію, магнію або кальцію змінює проникненість зябер, і здатний спричинювати масову загибель риб. Спалахи "цвітіння" Prymnesium зареєстровані у Болгарії, Ізраїлі, Данії, Голландії.
Викопні гаптофітові
Найдавніші рештки гаптофітових водоростей датуються віком біля 300 млн. років. Проте за показниками "молекулярного годинника", отриманими за результатами аналізу ядерного гену, що кодує 18S субодиницю цитолазматичної рибосомальної РНК, гетеротрофна клітина-господар гаптофітових виникла біля 850 млн років тому. Автотрофні примнезіофіти з'явилися пізніше - 320-260 млн років тому (за аналізами хлоропластних генів, що кодують велику субодиницю RUBISCO та 16S субодиницю хлоропластної рибосомальної РНК). Згідно до геологічного літопису, в юрському періоді (190-170 млн років тому) відбулося стрімке зростання різноманітності Haptophyta.
У крейдяному періоді гаптофітові стали провідною групою фітопланктону Світового океану. Майже всі відклади крейди того часу - це залишки коколітів представників Coccolithales. Цікаво, що майже до кінця 70-х років н.ст. панувало уявлення про те, що крейда - це залишки скелетів та панцирів фораменіфер, черепашкових амеб та радіолярій. Лише використання скануючого електронного мікроскопу дозволило з'ясувати справжнє походження крейди.
Наприкінці крейдяного періоду Haptophyta масово вимирають. Проте починаючи з середини третинного періоду (55-40 млн років тому) відділ знов переходить у стан біологічного прогресу, у якому перебуває і до сьогодення.
Відомості щодо викопних гаптофітових широко використовуються у геологічній практиці при визначенні віку осадових порід і при реконструкціях планетарного клімату у мезозойську та кайнозойську ери.
Місце Haptophyta у системі органічного світу
Перші гаптофітові були описані наприкінці ХІХ ст. На початку ХХ ст. А.Пашер розмістив їх серед золотистих водоростей як окремі роди класу Chrysophyceae. У 60-х роках електронно-мікроскопічні дані показали, що примнезіофіти мають кілька суттєво вімінних рис (зокрема, особливу джгутикоподібну структуру - гаптонему), і тому Т. Хрістенсен у 1962 виділив їх із Chrysophyceae в новий окремий клас Haptophyceae. У 1976 р. Д.Гібберд запропонував для гаптофіцієвих нову назву Prymnesiophyceae як узгоджену з вимогами Кодексу ботанічної номенклатури. У 80-х роках накопичився великий обсяг фактичного матеріалу, який не тільки підтверджував відокремленість примнезіофіцієвих від золотистих водоростей, але й свідчив про їх більш високий таксономічний статус. Тому наприкінці 80-х років гаптофіцієві (примнезіофіцієві) водорості було виділено у самостійний відділ Haptophyta (Prymnesiophyta).
На початку 90-х років гаптофітові водорості на підставі цитологічних даних (в першу чергу, за наявністю мітохондрій з трубчастими кристами) розглядали як примітивну бічну гілку Хроміст, яка має певні риси схожості також з криптофітовими водоростями. Починаючи з середини 90-х років численні молекулярно-філогенетичні реконструкції показують, що Haptophyta - це своєрідна перехідна філа, що знаходиться при основі царства Platycristates та підцарства Plantae. За ядерним геномом гаптофітові подібні до інших платикристат, за мітохондріальним геномом зберігають риси схожості з тубулокристатами. Здатності до фотосинтезу Haptophyta набувають вже після розходження платикристат та тубулокристат, внаслідок симбіозу з червоними водоростями, які і трансформуються у хлоропласт гаптофітових.