Альгологія - Костіков І.Ю. - 2009-2013
Розділ 21. Криптофітові водорості – Cryptophyta
Відділ Cryptophyta нараховує близько 200 видів одноклітинних мікроскопічних монадних водоростей, поширених як у континентальних водоймах, так і в морях. До початку 70-х років криптофітові водорості разом з динофітовими та рафідофітовими об'єднували у відділі Pyrrophyta на підставі подібності забарвлення, монадної будови, наявності глотки та еджективних структур. У 70-і роки криптофітові були виділені у самостійний відділ, який на підставі ряду цитологічних ознак розглядали як особливу філу примітивних представників рослинного світу. Після дослідження ядерного, хлоропластного та мітохондріального геномів, а також особливої структури, що містить ДНК - нуклеоморфа, вияснилось, що Cryptophyta споріднені з групою організмів, які мають мітохондрії з пластинчастими кристами, зокрема, з глаукоцистофітовими, червоними та зеленими водоростями.
Сьогодні Cryptophyta розглядають як відділ, що несе прямі докази походження пластид з хлоропластним ендоплазматичним ретикулюмом шляхом симбіозу типу "гетеротрофний евкаріот + фотоавтотрофний евкаріот".
Систематичні ознаки відділу
Пігменти та запасні поживні речовини
Для фотоавтотрофних криптофітових характерні хлорофіли а та с, α- та ε-каротини (Р-каротин, властивий переважній більшості інших відділів водоростей, у представників Cryptophyta не виявлений), фікобілінові пігменти - фікоціанін та фікоеритрин, а також дві групи ксантофілів: ксантофіли лютеїнового ряду (лютеїн, віолаксантин, зеаксантин), та кілька специфічних ксантофілів (алоксантин, крококсантин, монадоксантин). На відміну від синьозелених, глаукоцистофітових та червоних водоростей, у криптофітових фікобілінові пігменти знаходяться не у фікобілісомах, а безпосередньо на мембранах тилакоїдів з їх внутрішнього боку.
Кількісні співвідношення хлорофілів, різних фікобілінів та специфічних ксантофілів варіюють у межах різних родів у широкому діапазоні. Це обумовлює велику різноманітність забарвлення криптофітових водоростей. Так, хлоропласти можуть бути червоними, блакитними, жовтими, коричневими, зеленими. Є також види, що не мають хлоропластів - вони безбарвні і живляться виключно гетеротрофно.
Основним продуктом асиміляції у Cryptophyta є крохмаль. Він не відкладається у хлоропласті, як у зелених водоростей, або у цитоплазмі, як у глаукоцистофітових, а накопичується у перипластидному просторі, досить часто - поблизу піреноїда. Додатковим асимілятом є олія.
Цитологічні ознаки
Клітини криптофітових звичайно дорзовентральні, на передньому кінці косо зрізані, мають глибоку чи невелику глотку, два гетероконтні джгутики, що відходять від дорзального боку глоткового отвору. При основі глотки у прісноводних видів є одна-дві, рідше п'ять-вісім скоротливих вакуолей. На вентральному боці розташовується вузька поздовжня борозенка.
Клітинні покриви високоспецифічні, відомі під назвою перипласт. Він складається з плазмалеми, яка з внутрішнього боку несе прямокутні чи полігональні білкові платівки. Останні прикріплюються до плазмалеми за допомогою білкових шипів. Зовнішній бік плазмалеми може бути голим, проте частіше на ньому є шар зовнішніх білкових платівок, що за формою повторюють внутрішні, або шар субмікроскопічних лусок, що прикріплюються до плазмалеми за допомогою фібрилярного матеріалу (рис. 21.1). Прямокутні платівки, що утворюють правильні ряди, звичайно надають клітинним покривам криптофітових характерної поздовжньої посмугованості.
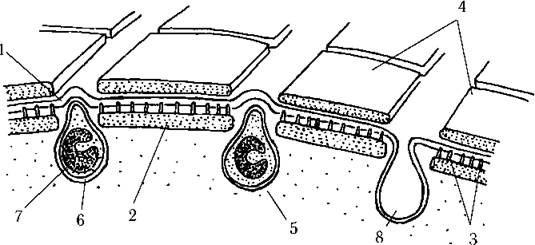
Рис. 21.1. Будова перипласта криптофітових водоростей: 1 - плазмалема, 2 - внутрішня білкова платівка, 3 - білкові шипи, що прикріплюють внутрішню платівку до плазмалеми, 4 - зовнішня платівка, 5 - трихоциста, 6 - мембранна капсула, 7 - згорнута стрічка трихоцисти, 8 - мембранна капсула трихоцисти після відстрілювання білкової стрічки (схематизовано за Lucas, 1970; Hausmann, 1979; Kugrens, Lee, 1987).
Під перипластом знаходяться трихоцисти. Великі трихоцисти звичайно вистилають глотку, дрібні розсіяні під усією поверхнею клітини. Трихоциста має вигляд мембранної капсули, всередині якої лежить щільно згорнута стрічка. Внаслідок подразнення плазмалеми, стрічка розгортається і вистрілює назовні, надаючи клітині реактивного поштовху. В результаті клітина може "стрибнути" вперед, раптово уповільнити рух або різко змінити його напрямок. Утворюються трихоцисти у пухирцях комплексу Гольджі.
Ядерний апарат евкаріотичний. Ядро звичайно розташовується в задній частині клітини, має досить велике ядерце. Зовнішня ядерна мембрана безпосередньо переходить у зовнішню мембрану хлоропласта (рис. 21.2). Центриолі відсутні, а їх функцію виконують базальні тіла джгутиків та ризопласти, до яких на початку мітозу мігрує ядро. Хромосоми в інтерфазі знаходяться у конденсованому стані, їх кількість коливається в межах 40-200.
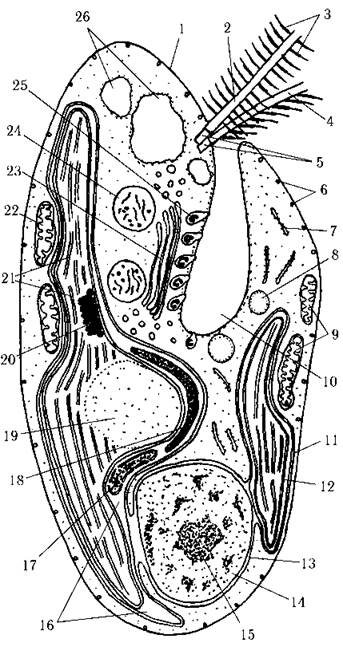
Рис. 21.2. Будова клітини криптофітової водорості: 1 - перипласт, 2 - довший пірчастий джгутик, 3 - мастигонема, 4 - коротший гребінчастий джгутик, 5 - базальні тіла джгутиків, 6 - дрібні трихоцисти, 7 - ендоплазматичний ретикулюм, 8 - краплина олії, 9 - вентральна мітохондрія, 10 - глотка, 11 - двомембранний хлоропластний ендоплазматичний ретикулюм, 12 — дві внутрішні мембрани оболонки хлоропласта, 13 - ядерна оболонка, 14 - ядро, 15 - ядерце, 16 - перипластидний простір, 17 - нуклеоморф, 18 - крохмаль, 19 - піреноїд, 20 - стигма, 21 - двотилакоїдні ламели, 22 - дорзальна мітохондрія, 23 - комплекс Гольджі, 24 - тільце Мопа, 25 - великі глоткові трихоцисти, 26 - скоротливі вакуолі (схематизовано за Lucas, 1970, 1982; Santore, Leedale , 1985).
Мітоз відкритий, певною мірою нагадує поділ ядра у примнезіофітових. Так, у метафазі хромосоми утворюють щільну платівку, індивідуальні хромосоми не розрізняються. Платівка має кілька каналів, через які проходять пучки мікротрубочок веретена. Мікротрубочки на полюсах не конвергують, а лишаються паралельними і закінчуються біля цистерн ендоплазматичного ретикулюму (рис. 21.3).
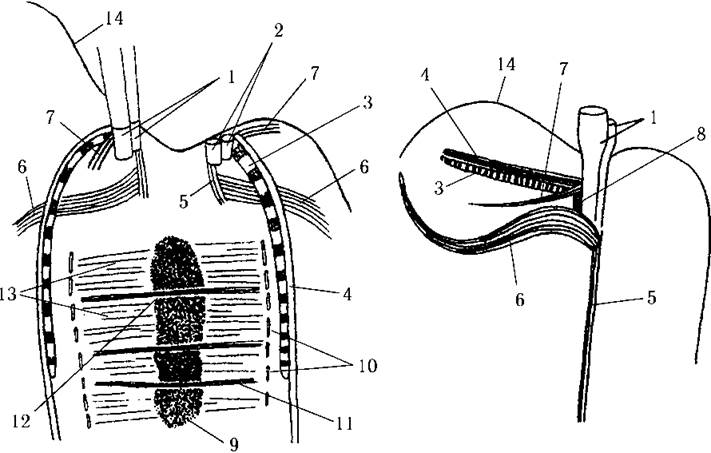
Рис. 21.3. Мітоз (ліворуч) та коренева система джгутиків (праворуч) у криптофітових водоростей: 1 - базальні тіла джгутиків, 2 - реплікована пара базальних тіл, 3 - ризопласт, 4 - мікротубулярний ризопластний корінь, 5 - ризостиль, 6, 7 - прості мікротубулярні корені, 8 - гладенький мікрофібрилярний корінь, 9 - хроматинова платівка, 10 - цистерни ендоплазматичного ретикулюму, 11 - пучок мікротрубочок, що проходить крізь хроматинову платівку, 12 - канал у хроматиновій платівці, 13 - поодинокі мікротрубочки веретена, 14 - перипласт (схематизовано за Oakley, 1978 та Седовою, 1996).
Фотосинтетичний апарат. Звичайно у клітині є один-два великі пристінні хлоропласти, вкриті чотиримембранною оболонкою. Дві зовнішні мембрани утворюють хлоропластну ендоплазматичну сітку. Зовнішня мембрана хлоропластного ендоплазматичного ретикулюму безпосередньо переходить у зовнішню мембрану ядра. Перипластидний простір між двома зовнішніми та двома внутрішніми мембранами хлоропласта містить крохмаль, 80S рибосоми та особливу органелу - нуклеоморф. Хоча перипластидний простір криптофітових і має свій власний білок-синтезуючий апарат, проте перипластидний ретикулюм відсутній.
Крохмаль відкладається у вигляді окремих зерняток або утворює суцільні крохмальні платівки. Останнє спостерігається лише у видів, що мають піреноїд.
Нуклеоморф - це органела, відмежована від перипластидного простору, в якому вона знаходиться, подвійною мембраною, яка, подібно до ядерної, має пори та містить ДНК й РНК. Нуклеоморф розташовується або поблизу піреноїда, або вільно лежить у перипластидному просторі. ДНК нуклеоморфа складається приблизно з 600 тис. пар нуклеотидів, організована у три хромосоми і не пов"язана з гістонами. Кожна хромосома нуклеоморфа містить гени, що кодують рибосомальну РНК та принаймні кілька білків. Поділ нуклеоморфа передує поділу ядра. Аналіз генів нуклеоморфу, що кодують 5S та 18S субодиниці РНК перипластидних рибосом показав, що геном нуклеоморфа близький до ядерного геному червоних водоростей.
Під внутрішніми мембранами оболонки хлоропласта знаходяться тилакоїди, піреноїд, вічко, власна хлоропластна ДНК, хлоропластні 70S рибосоми. Оперезуюча ламела відсутня. Тилакоїди зібрані по два. Загалом у хлоропласті спостерігається до кількох десятків таких двотилакоїдних ламел. На внутрішніх боках кожної пари безпосередньо на мембранах локалізовано фікобілінові пігменти, які, проте, не організовані у фікобілісоми. Піреноїд є у багатьох криптофітових. Він звичайно великий, напівзануреного типу, його вільна поверхня орієнтована до середини клітини. У зоні перипластидного простору, що безпосередньо прилягає до піреноїда, відкладається крохмаль. У деяких криптофітових у хлоропласті є вічко, що складається з кількох шарів пігментних глобул і прилягає до хлоропластної мембрани; положення вічка не зкоординовано із джгутиковим апаратом.
Хлоропластна ДНК розділена на кілька фрагментів, що знаходяться у різних частинах строми хлоропласта, тобто генофор є розсіяним.
Серед Cryptophyta є гетеротрофні види, що вторинно втратили фотосинтетичні пігменти або й хлоропласт, а також первинно гетеротрофні представники. Вторинні гетеротрофи - наприклад, з роду Chilomonas - зберігають редуковану пластиду - лейкопласт.
Мітохондріальний апарат звичайно представлений двома великими розгалуженими мітохондріями - дорзальною та вентральною. Обидві мають пластинчасті кристи і за допомогою мікротубулярного та мікрофібрилярного коренів з'єднуються з базальними тілами джгутиків. Мітохондріальна ДНК складається приблизно з 50 тис. пар нуклеотидних основ.
Джгутиковий апарат. Обидва джгутики у криптофітових звичайно спрямовані вперед На довшому джгутику мастигонеми розташовуються у два ряди, на коротшому вони однорядні. Мастигонеми мають розширену та звужену частини, і цим відрізняються від мастигонем усіх інших груп евкаріот. Синтезуються мастигонеми в ендоплазматичному ретикулюмі. Крім того, мастигонеми криптофітових вкриті органічними субмікроскопічними лусочками характерної форми. У перехідній зоні джгутики мають по дві поперечні платівки.
Коренева система джгутиків складається з чотирьох мікротубулярних та двох мікрофібрилярних коренів. До перших належать ризостиль, два простих кореня та корінь, пов'язаний з ризопластом. Мікрофібрилярні корені представлено поперечно-смугастим ризопластом та гладеньким коренем.
Ризостиль складається з 6-8 паралельних мікротрубочок, він спрямований до заднього кінця клітини, проходить поблизу ядра і, вірогідно, виконує опірну функцію. Перший простий мікротубулярний корінь побудований з чотирьох мікротрубочок і пов'язує базальні тіла джгутиків з дорзальною мітохондрією; другий корінь, утворений двома мікротрубочками, охоплює скоротливі вакуолі. Останній мікротубулярний корінь складається з чотирьох-шести мікротрубочок і щільно прилягає до ризопласта (див. рис. 21.3).
Поперечно-смугастий мікрофібрилярний корінь - ризопласт - побудований з білка центрина, він проходить безпосередньо під перипластом і закінчується поблизу ядра. На початку мітозу ризопласт разом з базальними тілами джгутиків стає центром організації мікротрубочок веретена. Гладенький мікрофібрилярний корінь з'єднується з вентральною мітохондрією.
Інші органели. Специфічними органелами криптофітових є пара т.з. тілець Мопа. Це досить великі, добре помітні одномембранні структури, що знаходяться поблизу глотки і містять переважно залишки трихоцист. Припускають, що тільця Мопа є гомологами лізосом і їх функція полягає в перетравленні надлишкових трихоцист. Звичайно між глоткою та тільцями Мопа розташовується один комплекс Гольджі.
Типи структури тіла
Майже всі криптофітові водорості мають монадний тип структури тіла. Переважна більшість видів здатна втрачати джгутики, одягатися шаром слизу і переходити у пальмелевидний стан. Одному роду - Bjornbergiella - властивий нитчастий тип структури.
Розмноження та життєві цикли
Основний спосіб розмноження Cryptophyta - це поділ клітин надвоє у поздовжньому напрямку в рухливому стані. Одна з дочірніх клітин отримує обидва материнські джгутики, друга утворює їх заново. Нестатеве розмноження за допомогою спеціалізованих клітин у криптофітових водоростей не виявлене.
Тривалий час вважалося, що статевий процес у криптофітових водоростей відсутній. У 1986 р. у Proteomonas sulcata був описаний диплогаплофазний життєвий цикл з гетероморфною зміною поколінь. Обидва покоління - гаплоїдне і диплоїдне - були представлені монадними клітинами і розмножувались поділом клітини надвоє. Диплоїдні клітини відрізнялися від гаплоїдних більшими розмірами. На жаль, спостерігати статевий процес та мейоз не вдалося.
У 1988 р. статевий процес гологамного типу був відкритий у Chroomonas acuta. Після злиття клітин відбувалася каріогамія і утворювалась диплоїдна планзигота. Проте місце проходження мейозу та тип життєвого циклу лишилися нез'ясованими. Таким чином, статеве розмноження у криптофітових існує, хоча його деталі лишаються недослідженими.
Основним типом життєвого циклу у переважної більшості криптофітових є цикломорфоз. У сприятливих умовах клітини розмножуються поділом, не припиняючи руху. Погіршення умов вегетації призводить до втрати джгутиків, виділення слизу і переходу до пальмелевидного стану, під час якого клітини зберігають здатність до поділу. При покращенні умов клітини відновлюють джгутики і повертаються до монадного стану. Наприкінці вегетаційного сезону криптофітові водорості можуть утворювати цисти: клітини втрачають джгутики, набувають сферичної форми, одягаються товстим шаром щільного слизу і переходять у стан спокою. Після закінчення періоду спокою циста проростає двома дочірніми монадними клітинами (рис. 21.4).
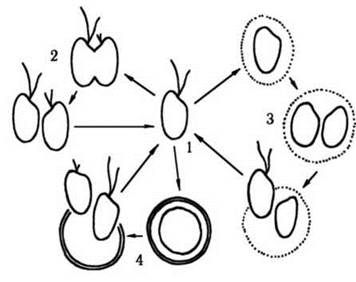
Рис. 21.4. Життєвий цикл криптофітової водорості: 1 - монадна вегетативна клітина, 2 - розмноження поділом у рухливому стані, 3 - пальмелевидний стан, 4 - циста та її проростання.
Система відділу
Всі Cryptophyta належать до одного класу - Cryptophyceae та одного порядку - Cryptomonadales. В основу поділу порядку на родини та роди покладено такі ознаки, як наявність розвиненої глотки, хлоропластів та лейкопластів, забарвлення клітин.
Так, глотка дуже редукована в межах родини Cryptochrysidaceae, характерними представниками якої є фотоавтотрофні водорості з бурими або червонуватими хлоропластами з роду Cryptochrysis. Добре розвинену глотку мають усі представники родини Cryptomonadaceae, в межах якої роди розрізняють, в першу чергу, за кольором. Так, роду Cryptomonas властиві жовті або коричневі хлоропласти, Rhodomonas - червоні, Chroomonas - сині. У видів роду Chilomonas хлоропласти редуковані, пластиди зберігаються у вигляді безбарвних лейкопластів. Єдиний рід з родини Cyathomonadaceae - Cyathomonas - позбавлений будь-яких пластид і за системами різних авторів розглядається або як найпримітивніший представник криптофітових, або як вершина еволюції даного відділу (рис. 21.5).
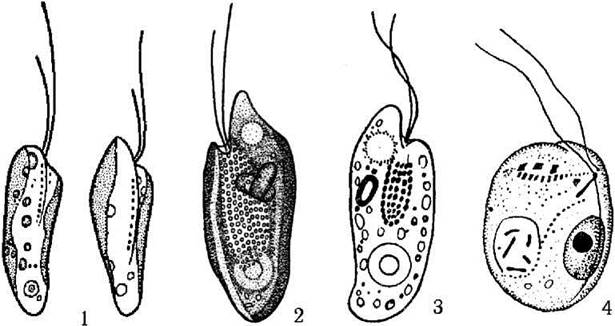
Рис. 21.5. Деякі представники криптофітових водоростей: 1 - Cryptochrysis commutata, 2 - Cryptomonas skujae, 3 - Chilomonas paramaecium, 4 - Cyathomonas truncata (за Матвієнко, Литвиненко, 1977).
Цікаво, що електронно-мікроскопічні та молекулярно-біологічні дослідження показали, що частина безбарвних водоростей, які традиційно відносили до криптофітових (зокрема, родина Katablepharidaceae), має не пластинчасті, а трубчасті мітохондріальні кристи, і тому повинна бути вилучена з Cryptophyta і вміщена у новий самостійний таксон рангу типу/відділу або класу.
Поширення, екологія, значення
Криптофітові водорості мешкають переважно у прісних водоймах або в морях. Винятки нечисленні - наприклад, Cryptomonas salina та C.stigmatica є галофілами і часто зустрічаються у планктоні солоних озер при концентрації солей 40-90 г/л; Cryptomonas cryophila та C. frigoris - кріофіли, які розвиваються на поверхні снігу в горах та викликають його червоне та зелене "цвітіння".
У прісних водоймах більшість криптофітових водоростей мешкає у товщі води чистих (наприклад, види роду Chroomonas), рідше - забруднених водойм (Cryptomonas ovata, C. erosa, і особливо - безбарвні представники роду Chilomonas).
Морські криптофітові звичайно зустрічаються у супраліторальних ваннах та прибережних калюжах. Відомі також кілька видів, що розвиваються у планктонних угрупуваннях відкритого океану, зокрема - у Північному морі.
Криптофітові водорості є переважно холодолюбними або евритермними організмами, нейтрофілами, рідше - ацидофілами. Відомі представники, що мешкають у сірководневому середовищі - серед гниючих рослинних решток та у глибинних шарах озер.
Деякі види Cryptophyta використовуються як індикатори забруднення водойм органічними речовинами, вони беруть участь у самоочищенні природних водойм та доочистки стічних вод у біологічних ставках.
Місце в системі органічного світу
За комплексом цитологічних ознак (зокрема, за наявністю специфічних двочленних мастигонем, субмікроскопічних лусочок на джгутиках, мітозу з утворенням щільної метафазної платівки, ядра, що з'єднується з хлоропластом зовнішньою мембраною хлоропластної ендоплазматичної сітки) криптофітові водорості подібні до гаптофітових. За хімічним складом асимілятів, мітохондріальними пластинчастими кристами простежуються риси подібності до зелених та глаукоцистофітових водоростей. Наявність фікобілінів споріднює криптофітових з червоними водоростями та глаукоцистофітовими. Проте деякі суттєві ознаки чітко відрізняють Cryptophyta від переважної більшості інших відділів водоростей. Це, в першу чергу, стосується нуклеоморфа.
За особливостями ядерного геному криптофітові займають проміжне місце між гаптофітовими, з одного боку, і водоростями з первинно- симбіотичними пластидами - глаукоцистофітовими, зеленими та червоними водоростями, з другого, мітохондріальний геном також нагадує мітохондріальні геноми зелених, червоних та глаукоцистофітових водоростей. Хлоропластний геном криптофітових найбільше споріднений з хлоропластними геномами червоних водоростей, а геном нуклеоморфа - з ядерним геномом останніх.
Тому вважають, що гетеротрофні пращури Cryptophyta мали спільного предка з гетеротрофними Prymnesiophyta, і дали початок двом гілкам. Перша стала "батьківською" для водоростей з двомембранними пластидами - Glaucocystophyta, Chlorophyta, Rhodophyta. Друга гілка привела до гетеротрофних криптофітових. Останні набули здатності до фотосинтезу порівняно недавно, внаслідок вторинного симбіозу з клітинами червоних водоростей. Ядро червоної водорості, що дала початок хлоропласту криптофітових, збереглось у вигляді нуклеоморфа.