Альгологія - Костіков І.Ю. - 2009-2013
Розділ 22. Глаукоцистофітові водорості – Glaucocystophyta
Відділ Glaucocystophyta нараховує біля 10 видів вільноіснуючих мікроскопічних одноклітинних водоростей, поширених у прісних континентальних водоймах. Викопні рештки глаукоцистофітових поки що не знайдені.
Всі відомі види глаукоцистофітових є фотоавтотрофами. Найцікавіша та найяскравіша особливість представників відділу - це наявність унікального фотосинтетичного апарату, представленого цианелами - видозміненими симбіотичними синьозеленими водоростями, які втратили здатність до самостійного існування поза межами клітини-господаря. Через це глаукоцистофітові водорості сьогодні навіть називають "прикладом симбіогенетичної еволюції у дії". Саме з вивчення геномів клітини-господаря та цианели було розпочато дослідження у галузі філогенії пластид на молекулярно-біологічному рівні.
За будовою інших органел глаукосистофітові мають суттєві риси схожості з криптофітовими, зеленими та гаптофітовими водоростями.
Систематичні ознаки відділу
Пігменти та запасні поживні речовини
Склад пігментів глаукоцистофітових практично такий самий, як і у синьозелених водоростей: наявні хлорофіл а, Р- каротин, фікобілінові пігменти - фікоцианін та аллофікоцианін. На відміну від Cyanophyta, у представників Glaucocystophyta поки що достовірно не знайдений фікоеритрин. Завдяки великій кількості фікобілінів цианели глаукоцистофітових забарвлені у блакитно-зелений колір. З ксантофілів виявлено β-криптоксантин, характерний також для криптофітів.
Продуктом асиміляції у Glaucocystophyta є звичайний крохмаль. Крохмаль глаукоцистофітових, на відміну від крохмалю зелених водоростей та вищих рослин, завжди відкладається поза межами фотосинтетичного апарату.
Цитологічні ознаки
Клітинні покриви глаукоцистофітових хоча і називають пелікулою, проте вони більше нагадують амфієсму динофітів: під плазмалемою розташовуються сплощені мембранні пухирці - лакуни, всередині яких можуть знаходитись тонкі платівки. Лакуни з боку цитоплазми підстилаються мікротрубочками (рис. 22.1). Наявність шару мікротрубочок відрізняє покриви глаукоцистофітових від типової амфієсми, а наявність лакун замість білкових стрічок - від пелікули евгленофітів.
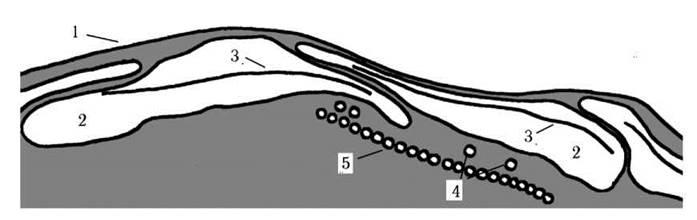
Рис. 22.1. Будова пелікули глаукоцистофітових водоростей: 1 - плазмалема, 2 - лакуна, відокремлена від цитоплазми власною мембраною, 3 - лакунарна платівка, 4 - підстилючі мікротрубочки, 5 - мікротрубочки джгутикового кореню (за Kies, 1989).
У деяких представників (наприклад, у роді Cyanoptyche) з зовнішнього боку пелікули спостергається також клітинна оболонка, утворена шаром сотоподібно розміщених субодиниць, над якими розташовується кілька шарів мікрофібрил поки що нез'ясованої природи.
Ядерний апарат типово еукаріотичний. Ядерна оболонка складається з двох мембран. Всередині ядра міститься велике ядерце. Ядерна ДНК пов’язана з білками-гістонами і має хромосомну організацію. Цікавою особливістю ядра глаукоцистофітових є наявність в ньому пучків мікрофіламентів. Крім глаукоцистофітових, подібні ядерні мікрофіламенти виявлено у деяких червоних водоростей.
За результатами аналізів окремих генів (в першу чергу, гену, що кодує 18S субодиницю рибосомальної РНК) виявлено, що ядерний геном глаукоцистофітових найбільш близький до ядерних геномів криптофітових, червоних та зелених водоростей, а також досить близький до геному гаптофітових водоростей.
Мітохондрії глаукоцистофітових розгалужені, на зрізах звичайно виглядають як численні невеликі пухирці з пластинчастими кристами (рис. 22.2), і можуть розглядатися як мітохондріон. За цитологічними даними, зокрема, за будовою крист, мітохондріальна система глаукоцистофітових є досить схожою з мітохондріонами інших платикристат. На жаль, мітохондріальний геном глаукоцистофітових поки що лишається не дослідженим.
Фотосинтетичний апарат Glaucocystophyta унікальний, він не має аналогів серед еукаріотичних водоростей і являє собою цианели. У літературі можна зустріти дві точки зору на природу цианел: їх вважають або попередниками справжніх хлоропластів еукаріот - симбіотичними синьозеленими водоростями, що втратили здатність до розвитку поза межами клітини-господаря, проте ще зберегли певні риси індивідуального організму, або ж як дуже примітивні хлоропласти. Звичайно у клітині міститься від двох (рід Cyanophora) до багатьох (роди Glaucocystis, Cyanoptiche) цианел (рис. 22.2: а,б; 22.3).
Кожна цианела відмежована від цитоплазми однією цитоплазматичною мембраною. Під цією мембраною знаходиться тонкий електронно-щільний шар, утворений пептидогліканом (муреіном) (рис. 22.2: в,г). З внутрішнього боку цього шару розташовується ще одна мембрана. Наявність у цианел шару муреіну між зовнішньою та внутрішньою мембранами чітко відрізняє фотосинтетичний апарат глаукоцистофітових від типового хлоропласту.
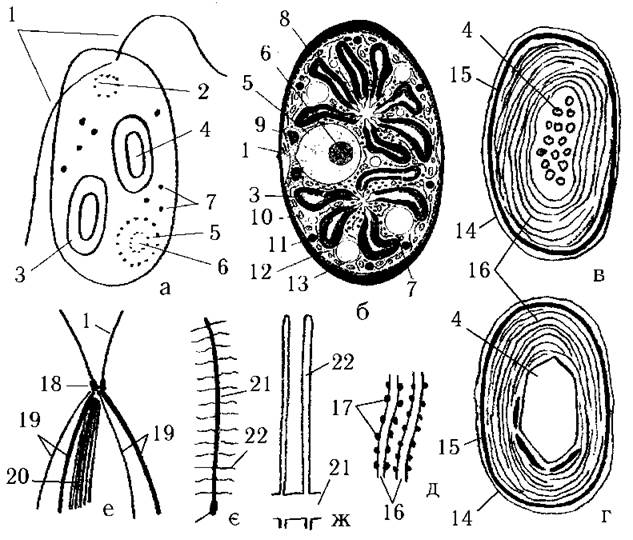
Рис. 22.2. Глаукоцистофітові водорості: а - зовнішній вигляд Cyanophora paradoxa в оптичному мікроскопі; б - схема будови Glaucocystis; в,г - будова цианел Cyanophora paradoxa (в) та Cyanoptiche gloeocystis(г); д - фікобілісоми на мембранах тилакоїдів; е-ж - схема кореневої системи джгутиків (е), джгутикової поверхні (є) та морфологія мастигонем (ж) глаукоцистофітів. 1 - джгутики, 2 - пульсуюча вакуоль, 3 - цианела, 4 - карбоксисома, 5 - ядро, 6 - ядерце, 7 - гранули крохмалю, 8 - клітинна оболонка, 9 - комплекс Гольджі, 10 - мітохондрія, 11 - краплина олії, 12 - ендоплазматичний ретикулюм, 13 - вакуоль з клітинним соком, 14 - зовнішня мембрана цианели, 15 - муреінова оболонка цианели, 16 - тилакоїди, 17 - фікобілісоми, 18 - базальне тіло джгутика, 19 - мікротубулярні джгутикові корені, 20 - багатошарова структура, 21 - аксонема, 22 - мастигонема (схематизовано за Kies, 1979, 1989; O'Kelly, 1992).
Всередині цианели концентричними колами розташовуються поодинокі тилакоїди. Поверхня мембран тилакоїдів несе на собі числені фікобілісоми, подібні до фікобілісом синьозелених та червоних водоростей. У центральній частині цианели розміщується одна велика або багато дрібних карбоксисом, утворених рибульозо-дифосфат-карбоксилазою. Ці карбоксисоми є повними аналогами поліедральних тілець синьозелених водоростей.
Цианели мають свою власну ДНК, яка замкнена в кільце та не пов'язана з гістонами. Сьогодні геном цианел одного з представників глаукоцистофітових - Cyanophora paradoxa - повністю розшифровано; також досить детально досліджено геном іншого глаукоцистофіта - Glaucocystis nostochinearum. Встановлено, що геном цианел містить приблизно в 10 разів менш нуклеотидів, ніж геном синьозелених водоростей (біля 125 тис. пар нуклеотидів замість 1.21.5 млн.). За майже всіма дослідженими генами цианели безпосередньо виводяться від синьозелених водоростей. Проте також встановлено, що цианели є "сестрами", а не пращурами вихідних типів хлоропластів еукаріот.
Джгутиковий апарат. Монадні стадії глаукоцистофітових дводжгутикові, дорзовентральні, можуть бути представлені як вегетативними клітинами (Cyanophora), так і зооспорами (Glaucocystis, Cyanoptyche). Джгутики гетероконтні та гетеродинамічні, виходять з субапікального заглиблення, яке є початком поздовжньої борозенки. Коротший джгутик направлений вперед і є рушійним. Довший джгутик лежить у поздовжній борозенці і виконує функцію стерна. Біля основи джгутиків розташовується одна пульсуюча вакуоль. У зооспор біля базальних тіл розміщується кілька диктіосом, які (принаймі, у Cyanoptyche) приймають участь у побудові клітинної оболонки після припинення руху клітини та прикріплення її до субстрату. Стигми у глаукоцистофітових не виявлено.
Кожний джгутик має два ряди простих, не тричастних мастигонем, і цим нагадує джгутики криптофітових та деяких зелених водоростей (рис. 22.2: є,ж).
Коренева система джгутиків складається з чотирьох хрестоподібно розміщених мікротрубочкових коренів та мікротубулярної багатошарової структури (рис. 22.2: е). До складу кожного кореня входить біля 30 мікротрубочок. Два корені з чотирьох з'єднуються з багатошаровою структурою. Багатошарова структура з зовнішніх боків несе електронно-щільні шари, між якими знаходиться шар мікротрубочок та шар паралельних платівок нез'ясованої природи. Самі ж базальні тіла поєднані між собою фібрилярною зв’язкою.
Плани будови тіла
Хоча сьогодні відомо лише чотири роди глаукоцистофітових, проте вони представляють два типи морфологічних структур: монадний та гемімонадний.
Так, для роду Cyanophora характерним є монадний тип структури (рис. 22.2, 22.3). Клітини цианофори поодинокі, колоній не утворюють.
Родам Cyanoptyche та Glaucocystis властива гемімонадна будова. Кожна клітина має пульсуючу вакуоль, а між клітинною оболонкою та пелікулою зберігаються сильно редуковані джгутики. Ці водорості об’єднані у глеокапсоподібні колонії, що складаються з 2-4-8 клітин і мають вигляд системи вкладених один в одного слизових міхурів. Відміни між Cyanoptyche та Glaucocystisполягають у кількості та формі цианел: у першого роду цианели дрібні, численні (до кількох десятків) і не утворюють зірчастих кластерів; у другого - цианели досить великі, порівняно нечисленні (8-15) і групуються в один-два кластери у вигляді зірок (рис. 22.3).
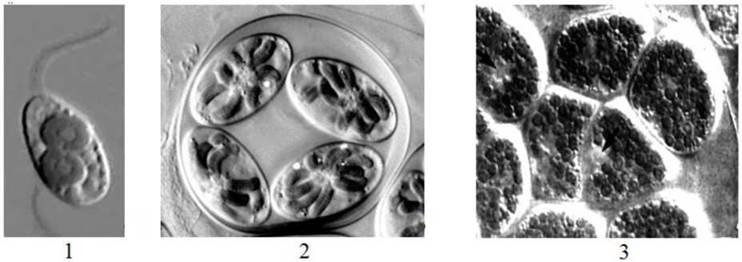
Рис. 22.3. Деякі глаукоцистофітові водорості: 1 - Cyanophora paradoxa; 2 - Glaucocystis nostochinearum; 3 - Cyanoptyche gloeocystis. Грілками позначено пульсуючі вакуолі (за Kies, 1989).
Розмноження та життєві цикли
Монадні глаукоцистофітові водорості розмножуються вегетативним поділом клітини надвоє у рухливому стані. Поділу клітини передує поділ цианел. Таким чином, поділи клітини і цианел є досить скоординованими.
Колоніальні представники з гемімонадною будовою (наприклад, Glaucocystis) можуть розмножуватись простим поділом клітин надвоє, шляхом фрагментації колоній, а також за допомогою зооспор. Зооспори глаукоцистофітових за своєю морфологією подібні до вегетативних клітин Cyanophora.
Статеве розмноження у глаукоцистофітових не виявлено і життєвий цикл являє собою цикломорфоз.
Система відділу
Відділ включає один клас - Glaucocystophyceae і два порядки - Cyanophorales та Glaucocystales. Перший порядок об'єднує види з монадним типом структури тіла і містить лише один рід - Cyanophora. До другого порядку входять представники з гемімонадною будовою — роди Glaucocystis, Cyanoptyche, Gloeochaete (рис. 22. 3).
Поширення, екологія та значення
Всі глаукоцистофітові є мешканцями континентальних прісних водойм. Види роду Cyanophora розвиваються переважно в планктоні ефемерних водоймах, забруднених органічною речовиною. Види родів Glaucocystis, Cyanoptyche та Gloeochaete є мешканцями перифітону заплавних водойм. Наприклад, одразу всі ці три роди було виявлено в одній й тій ж самій стариці р. Дунай в обростаннях вищих водних рослин Hippuris vulgaris та Utricularia minor.
Глаукоцистофітові є рідкісними водоростями. На території України знайдено два види з цього відділу - Cyanophora paradoxa та Glaucocystis nostochinearum.
У практичній діяльності людини Glaucocystophyta не використовуються. Проте вони мають надзвичайно важливе теоретичне значення у вирішенні питань філогенії водоростей та органічного світу в цілому.
Цианели глаукоцистофітових та походження хлоропластів
Дослідження глаукоцистофітових відіграли значну роль у з’ясуванні питання про походження та еволюцію хлоропласту. У зв’язку з цим цікавою є історія розвитку поглядів на природу цианел.
Перші глаукоцистофітові - монадна Cyanophora paradoxa та гемімонадна Cyanoptyche gloeocystis - були описані на початку нашого сторіччя як представники різних відділів: перший вид був віднесений до класу Cryptophyceae розформованого зараз відділу Pyrrophyta, а другий вид - до порядку Tetrasporales відділу Chlorophyta. Обидва види вважалися гетеротрофами, що утворюють стійкий симбіоз з якимись організмами, вірогідно - з синьозеленими водоростями. На той час визначити таксономічне положення ендосимбіонтів більш глибоко не вдалося, тому ці "організми" було запропоновано розглядати як синьозелені водорості нез’ясованого систематичного положення і називати цианелами.
У 70 - 80-х роках н.ст. перші монадні та гемімонадні глаукоцистофітові були досліджені за допомогою електронного мікроскопу. На основі єдиного плану будови джгутикових стадій та цианел, види глаукоцистофітових, які до цього часу розміщувались у різних відділах, у 1984 р. були об'єднані в єдиний клас - Glaucocystophyceae, і далі виділені в окремий відділ Glaucocystophyta (або Glaucophyta).
Електронно-мікроскопічні дослідження 70-х - початку 80-х років, з одного боку, підтвердили те, що цианели за своєю будовою дійсно надзвичайно схожі з синьозеленими водоростями. З іншого боку, результати показали що, на відміну від Cyanophyta, цианели мають редуковану клітинну оболонку, яка позбавлена зовішнього фібрилярного целюлозно-пектинового шару, хоча і зберігає тонкий муреіновий шар. Ці факти (отримані до того ж у час найбільш гострої полеміки про симбіотичне походження еукаріотичної клітини) викликали дискусію про природу цианел: чи є цианели симбіотичними синьозеленими водоростями, чи вони являють собою особливий примітивний тип пластид?
У 80-х роках було здійснено перші спроби знайти відповідь за допомогою молекулярно-біологічних досліджень геному цианел. В результаті було відкрито дві надзвичайно цікаві особливості глаукоцистофітових: по-перше, розмір геному цианел (кількість пар нуклеотидів у ДНК цианел) майже співпадає з розміром геному хлоропластів інших еукаріот, і в середньому є в 10 разів меншими від геному Cyanophyta. По-друге, у геномі цианел було відкрито дві ділянки т.з. інвертованих послідовностей нуклеотидів. Ці ділянки являли собою гени, що кодують хлоропластну рибосомальну РНК. Транскрипція цих фрагментів відбувається у зворотних напрямках (тобто інвертовано). Інвертовані послідовності, що кодують хлоропластну рибосомальну РНК було виявлено також у хлоропластній ДНК вищих рослин, проте не знайдено у синьозелених водоростей. Фактично, молекулярні дослідження підтвердили припущення про те, що цианели - це хлоропласт, а не симбіотичні синьозелені водорості. Крім того, дані про розмір геному цианел (зокрема, його значна редукція порівняно з геномом прокаріот) дозволили зрозуміти, чому цианели не вдається культивувати за межами клітини.
Визнання цианел особливим примітивнм типом пластид призвело до побудови простого філогенетичного ряду, що пояснював походження хлоропластів: вільноіснуюча синьозелена водорость вступає в симбіоз з гетеротрофною клітиною-господарем і внаслідок редукції геному трансформується в примітивну пластиду - цианелу. Подальша редукція цианели, пов'язана з повною втратою власної клітинної оболонки, веде до виникнення справжнього хлоропласту - хлоропласту червоних водоростей. Червоні водорості різними шляхами (зокрема, в процесі дарвіновської еволюції або через вторинні симбіози типу "гетеротрофний еукаріот - червона водорість") дають початок всім іншим відділам водоростей. Таким чином, глаукоцистофітові виступають найбільш примітивними еукаріотичними рослинами. Ця точка зору стала дуже популярною у середині-наприкінці 90-х років нашого сторіччя: наприклад, згідно з нею подано матеріал у найсучаснішому підручнику з альгології - "Algae. An introduction to phycology" (C. Van den Hoek, D.G. Mann, H.M. Jahns, 1995, Cambrige Univ. Press).
Перевірка гіпотези про примітивність глаукоцистофітових була здійснена методами молекулярної філогенії. Основним об'єктом аналізу було обрано ген, що кодує 16S субодиницю рибосомальної РНК у бактерій, синьозелених водоростей, у цианелах глаукоцистофітових та у хлоропластах еукаріотичних водоростей практично всіх відділів (пізніше аналізувались також і інші гени - разом більш десяти). Результати виявилися несподіваними (рис. 22.4).

Рис. 22.4. Схема філогенетичних зв'язків хлоропластів різних типів. 1 - примітивні синьозелені водорості, що походять від фотоавтотрофних аноксигенних бактерій; 2 - всі інші синьозелені водорості; 3 - синьозелена водорость, що походить від примітивних Cyanophyta і є пращуром хлоропластів - гіпотетична т.з. пропластида; 4-6 - хлоропласти, що утворилися внаслідок симбіозу пропластиди з гетеротрофними еукаріотами - т.з., первинно-симбіотичні пластиди (4 - цианели глаукоцистофітових; 5 - хлоропласти зелених водоростей, 6 - хлоропласти червоних водоростей); 7 - хлоропласти вищих рослин - деривати хлоропласту зелених водоростей; 8-9 - вторинно-симбіотичні хлоропласти, що утворюються після симбіозу зеленої водорості з різними гетеротрофними еукаріотами (8 - хлоропласти евгленофітових водоростей, 9 - хлоропласти хлорарахніофітів); 10-14 - вторинно-симбіотичні хлоропласти, що утворилися після симбіозу червоної водорості з різними гетеротрофними еукаріотами, та їх деривати (10 - хлоропласти примнезіофітів, 11 - криптофітів, 12 - водоростей - хроміст, 13 - переважної більшості динофітів, 14 - вторинно- редуковані пластиди апікомплекс).
Було встановлено, що примітивні синьозелені водорості, що походять від фотоавтотрофних бактерій, дали початок п'ятьом-семи гілкам. Всі ці гілки, за винятком однієї, об'єднують сучасні синьозелені водорості (включаючі прохлорофітів) і є тупиковими. Проте одна гілка веде безпосередньо до хлоропластів еукаріот. Вважають, що в межах цієї гілки була представлена синьозелена водорість, здатна вступати у симбіоз з різноманітними гетеротрофними еукаріотами - т.з. пропластида. Пропластида далі дала початок трьом типам пластид, які називають первинно-симбіотичними: цианели глаукоцистофітових, хлоропласту зелених водоростей (т.з. пластиді хлорофітного типу) та хлоропласту червоних водоростей (пластиді родофітного типу). Всі інші типи хлоропластів є або більш пізніми дериватами цих первинних пластид (наприклад, хлоропласти вищих рослин), або виникли внаслідок вторинних симбіозів первинно-пластидних водоростей з різними гетеротрофними еукаріотами.
Таким чином, було встановлено, що цианели є не проміжним типом фотосинтетичного апарату у ряду "фотоавтотрофні прокаріоти - справжні хлоропласти", а, навпаки, тупіковим варіантом первинно-симбіотичних пластид.
Сьогодні глаукоцистофітові знов стали "об’єктом особливої уваги" систематиків та філогенетиків у зв'язку з новою дискусією: чи виникли три типи первинних пластид внаслідок лише одного акту симбіозу пропластиди з гетеротрофним еукаріотом, чи мали місце три різних симбіотичних акта з різними клітинами-господарями, що, відповідно, дало початок трьом різним відділам первинно-пластидних еукаріот. Зараз почали з'являтися факти на користь першої гіпотези, але до більш-менш обгрунтованого висновку поки що далеко.
Місце Glaucocystophyta у системі органічного світу
Сьогодні місце глаукоцистофітових у системі органічного світу визначено досить чітко. За будовою мітохондрій глаукоцистофітові є типовими платикристатами і, таким чином, споріднені з криптофітовими, червоними та зеленими водоростями, з вищими рослинами, а також з багатоклітииними тваринами та справжніми грибами.
За фотосинтетичним апаратом глаукоцистофітові є рослинами з первинно-симбіотичними пластидами - цианелами. Цитологічною ознакою первинно-симбіотичного характеру пластид Glaucocystophyta є наявність у складі хлоропластної оболонки лише двох мембран, а апоморфною ознакою - наявність муреїнового шару між цими мембранами. Цианели глаукоцистофітових та хлоропласти зелених і червоних водоростей утворюють єдину кладу, що виводиться від синьозелених водоростей.
За продуктами асиміляції, системою органел нуклеоцитоплазми, джгутиковим апаратом (особливо - за будовою мастигонем та багатошарової структури) глаукоцистофітові найбільш близькі до зелених, і в меншому ступені - до криптофітових водоростей. Спорідненість цих відділів підтверджено також результатами досліджень більш ніж десяти генів, що кодуються ядерною ДНК.
Весь цей комплекс ознак свідчить про те, що глаукоцистофітові та зелені водорості походять від спільного предка. На "роль" такого предка найбільш обґрунтовано претендують первинно-гетеротрофні криптофітові водорості або їх попередники.