Альгологія - Костіков І.Ю. - 2009-2013
Розділ 23. Червоні водорості – Rhodophyta
Червоні водорості, або багрянки - Rhodophyta - нараховують біля 5000 видів переважно багатоклітинних макроскопічних водоростей. Невелика частина видів представлена мікроскопічними формами. Майже всі червоні водорості є мешканцями бентосних та перифітонних морських угрупувань, і лише як виняток деякі види зустрічаються у прісних водоймах та у ґрунтах.
Викопні рештки Rhodophyta відомі з раннього палеозою (кембрій, 600 млн. років тому). За палеоальгологічними даними вважають, що в ордовику (500-400 млн. років тому) у відділі виникли гетеротрихальні форми. В середньому та пізньому палеозої (400-230 млн. років тому) Rhodophyta займали провідне положення в морях, на початку мезозою пережили період катастрофічного вимирання, внаслідок якого зникли майже всі види кам'яновугільної флори. У крейдяному періоді (біля 100 млн. років тому) відділ знов перейшов у стан біологічного прогресу і прогресує до теперішнього часу.
Червоні водорості належать до пластинчастокристних рослин, що мають пластиди первинно симбіотичного типу. Сестринськими по відношенню до Rhodophyta є відділи Glaucocystophyta та Chlorophyta.
Систематичні ознаки відділу
Пігменти та запасні поживні речовини
Клітини червоних водоростей забарвлені переважно у червоний колір, значно рідше забарвлення може бути сірим або синьо-зеленим (як правило, у водоростей, що мешкають у прісних водоймах або у ґрунтах). Забарвлення визначається, в першу чергу, наявністю хлорофілу а та фікобілінових пігментів - фікоцианіну, аллофікоцианіну та фікоеритрину. Звичайно фікоеритрин переважає над двома іншими фікобілінами. Каротиноїди представлені α- та β-каротинами, і двома групами ксантофілів - по-перше, специфічними ксантофілами (криптоксантином, тараксантином), по-друге, ксантофілами лютеїнового ряду - зеаксантином та лютеїном (а у деяких видів - ще й антероксантином та віолоксантином).
Основний продукт асиміляції - полісахарид багрянковий крохмаль - являє собою α-1,4 глюкан. Він близький до амілопектину та глікогену. Від розчину Люголю багрянковий крохмаль забарвлюється у жовто-бурий колір, далі стає червоним. У примітивних бангієвих водоростей продуктом асиміляції є справжній глікоген. У деяких Rhodophyta спостерігаються особливі секреторні залозисті клітини, що заповнені йодистими та бромистими сполуками, та клітини з іридіюючими протеїновими тільцями.
Додатковими запасними поживними речовинами можуть бути галактозид, флоризид, багатоатомні спирти (манніт), у незначній кількості - олія. Цікаво, що вуглевод флоризид виявлениий також у синьозелених та криптофітових водоростей. Основною функцією флоризиду є регуляція осмотичного тиску.
Всі продукти асиміляції у червоних водоростей відкладаються поза межами хлоропластів.
Цитологічні ознаки
Клітинні покриви представлено пектиновими та целюлозно-пектиновими оболонками. Оболонки звичайно тришарові. Зовнішній шар - кутикула - утворений, як правило, згущеною полімеризованою маннозою; середній шар - фікоколоїдами (агар-агаром, агароїдом, каррагеніном, які є сірковмісними гетерополісахаридами) та пектиновими речовинами. Внутрішній шар складається з целюлози (рис. 23.1). Примітивні бангієві водорості целюлозного шару позбавлені.
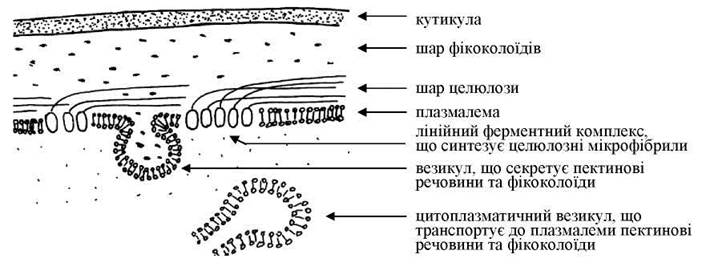
Рис. 23.1. Схема будови клітинної оболонки червоних водоростей.
Ріст середнього шару оболонки відбувається за рахунок діяльності комплексу Гольджі, внутрішнього - за рахунок синтезу мікрофібрил лінійними ферментними комплексами, розташованими у плазмалемі. У багатьох видів оболонки просякнуті вапном - кальцитом, арагонітом, карбонатами кальцію, магнію, заліза.
Зв'язок між клітинами відбувається за допомогою плазмадесм, що проходять крізь первинні та вторинні пори (рис. 23.2). Первинні пори утворюються внаслідок неповного змикання країв поперечної перегородки по закінченні поділу клітини. Первинні пори закриті спеціальними пробками.
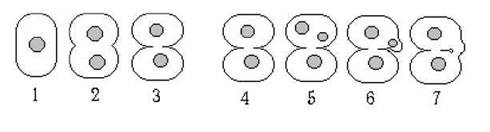
Рис. 23.2. Утворення первинних (1-3) та вторинних (4-7) пор у Rhodophyta.
В утворенні пробок безпосередню участь приймають цистерни ендоплазматичної сітки, завдяки яким формується білкова основа пробки, і інколи - її поверхнева мембрана. Крім того, пробки можуть мати додаткові внутрішні та зовнішні полісахаридні шари (рис. 23.3). У деяких представників зовнішній шар полісахаридів дуже товстий. Такі пробки добре помітні в оптичний мікроскоп.
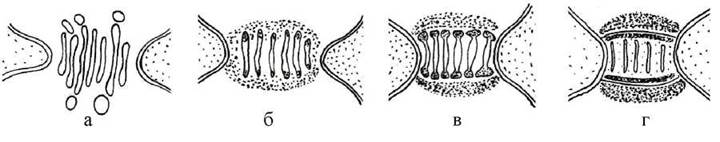
Рис. 23.3. Сема утворення пробки з поверхневою мембраною та додатковими шарами полісахаридів: а - у порі розташовуються цистерни ендоплазматичної сітки; б - навколо цистерн відкладається шар полісахаридів, у термінальних зонах цистерн накопичуються полісахариди майбутнього внутрішнього щару, між цистернами відкладається структурні білки, утворюючи основу пробки; в - термінальні ділянки цистерн відшнуровують везикули з полісахаридами; г - везикули з полісахаридами зливаються між собою, утворюючи поверхневу мембрану; білкова основа пробки, внутрішній та зовнішній полісахаридні шари ущільнюються.
Як наслідок, у червоних водоростей розрізняють сім типів пробок (рис. 23.4). Пробки першого, другого та третього типів не мають поверхневої мембрани, і відрізняються між собою за наявністю додаткових полісахаридних шарів. Так, на пробках першого типу такі шари відсутні, у другого типу є один тонкий додатковий шар; пробки третього типу мають два додаткові полісахаридні шари. Пробки без поверхневої мембрани характерні для представників класу бангєфіцієвих, а також для найпримітивніших представників класу флоридей.
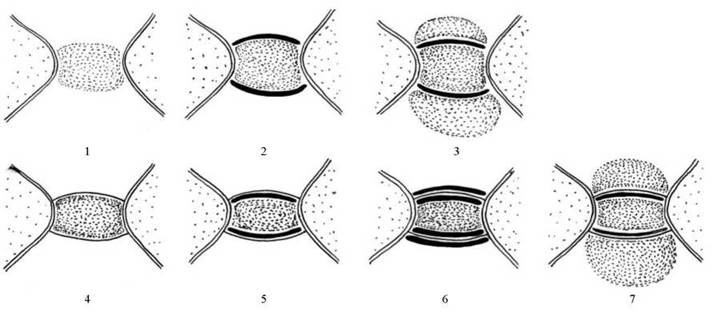
Рис. 23.4. Типи пробок у червоних водоростей. У верхньому ряду - пробки без поверхневої мембрани (перший, другий та третій типи), у нижньому ряду - з поверхневою мембраною (четвертий, п'ятий, шостий та сьомий типи).
У пробок четвертого-сьомого типів поверхневі мембрани наявні. Відміни в межах цих типів також пов'язані з додатковими полісахаридними шарами: в четвертому типі їх немає, у пробок п'ятого типу є один додатковий полісахаридний шар, розташований між мембраною та білковим тілом; у пробок шостого та сьомого типів є два додаткових полісахаридних шари, розмежовані поверхенвою мембраною, причому в пробках сьомого типу зовнішній шар дуже товстий і помітний в оптичний мікроскоп. Пробки з поверхневими мембранами характерні для тих представників класу Florideophyceae, які вважаються еволюційно більш продвинутими.
В цілому, тип пробок є тією ознакою, яка добре узгоджується з молекулярними даними щодо родинних зв'язків різних порядків Rhodophyta, і тому використовується як одна з важливих фенотипічних ознак на рівні класів та порядків.
Вторинні пори є наслідком особливого поділу: в клітині відбувається мітоз та утворюється дочірне ядро, далі це ядро мігрує у бічний випин оболонки; цей випин з'єднується з сусідньою клітиною, оболонка у місці контакту руйнується і з'являється вторинна пора. Дочірне ядро при цьому дегенерує.
Ядерний апарат типово еукаріотичний. Проте при мітозі спостерігаються деякі суттєві відміни від класичної схеми: мітоз є закритим, і ядерна оболонка у профазі не зникає, а перфорується. Навколо ядра у пізній профазі з частини ендоплазматичної сітки утворюється так званий перинуклеарний ендоплазматичний ретикулюм. Центриолі у червоних водоростей відсутні, а їх функцію виконують особливі білкові полярні кільця.
Після завершення поділу ядра відбувається цитокінез. При цьому плазмалема та внутрішній шар клітинної оболонки вростають у клітину, утоворюючи кільцеву борозну. Краї борозни зазвичай не змикаються, і в центрі нової поперечної перегородки залишається пора. Далі у зоні пори відкладається білковий матеріал і утворюється пробка.
Фотосинтетичний апарат. Хлоропласти червоних водоростей вкриті двомембранною оболонкою, тилакоїди поодинокі. На поверхні тилакоїдів зосереджено фікобілісоми, будова яких подібна до фікобілісом синьозелених водоростей.
Хлоропласти Rhodophyta умовно можна розділити на два типи за положенням у клітині, кількістю, наявністю піреноїдів та оперезуючого тилакоїду: а) поодинокі центральні зірчасті хлоропласти з голими піреноїдами; б) численні дрібні парієтальні дископодібні хлоропласти, в яких піреноїди відсутні. Хлоропласти першого типу притаманні переважно бангієфіцієвим водоростям, другого - представникам класу флорідей (рис. 23.5).
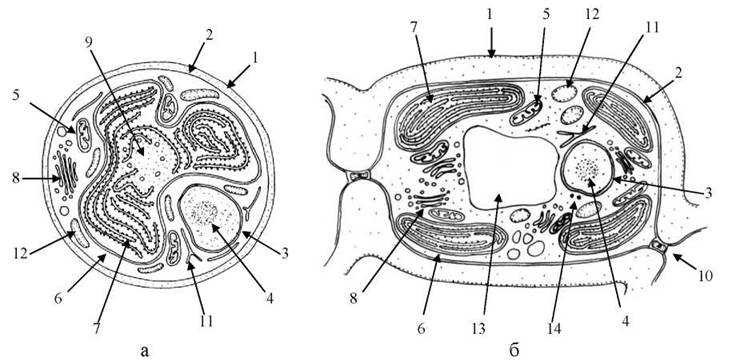
Рис. 23.5. Схеми будови клітин червоних водоростей класів Bangiophyceae (а) та Florideophyceae (б): 1 - клітинна оболонка; 2 - плазмалема; 3 - ядерна оболонка; 4 - ядерце; 5 - мітохондрія; 6 - оболонка пластиди; 7 - тилакоїд із фікобілісомами; 8 - комплекс Гольджі; 9 - піреноїд; 10 - пробка; 11 - ендоплазматична сітка; 12 - запасні полісахариди; 13 - вакуоля; 14 - полярні кільця.
За молекулярними даними, хлоропластна ДНК червоних водоростей найбільш споріднена, з одного боку, з ДНК нуклеоїду синьозелених водоростей, з іншого - з хлоропластною ДНК зелених та глаукоцистофітових водоростей. Це добре узгоджується з цитологічними даними про первино-симбіотичне походження пластид Rhodophyta.
Цікавим є те, що ген, який кодує RuBisCo (rbcL - ген), червоні водорості отримали від альфа-протеобактерій, внаслідок горизонтального його переносу, тоді як пластиди зелених водоростей набули цей ген від синьозелених водоростей. Припускають, що дивергенція гіпотетичної первинно-симбіотичної пластиди на хлоропласт (пластиду зелених водоростей) та родопласт (пластиду червоних) могла бути обумовлена саме різним походженням гену rbcL, який кодує ключовий фермент початкових етапів темнової фази фотосинтезу.
Джгутикові стадії у червоних водоростей повністю відсутні.
Плани будови талому
В межах відділу представлено кокоїдний, нитчастий та гетеротрихальний типи структур. Кокоїдні форми нечисленні, зустрічаються серед найпримітивніших бангієфіцієвих водоростей. Кокоїдні червоні водорості звичайно утворюють слизові колонії (рис. 23.6).
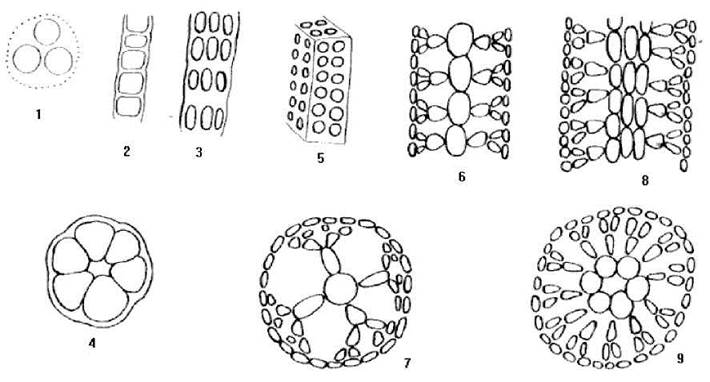
Рис.23.6. Типи морфологічних структур та плани будови таломів Rhodophyta: 1 - кокоїдний; 2 - нитчастий однорядний; 3, 4 - нитчастий багаторядний; 5 - пластинчастий; 6, 7 - гетеротрихальний одноосьовий; 8, 9 - гетеротрихальний багатоосьовий.
Водорості з нитчастим типом структури представлені в таксонах різного ступеню продвинутості. Таломи водоростей з нитчастим типом структури можуть мати ниткоподібну або пластинчасту форму, бути одно- або багаторядними. Деякі автори вважають, що пластинчасті багаторядні таломи слід розглядати як тканинні (наприклад, таломи Porphyra) (див. рис. 23.6).
Найбільш поширені в межах відділу представники з гетеротрихальним типом морфологічної будови тіла. Ріст таких водоростей відбувається переважно за рахунок поділу апікальної клітини. Бічні нитки-асимілятори нерідко щільно зростаються між собою, утворюючи псевдопаренхіматозні структури. В залежності від кількості опірних ниток гетеротрихальні таломи поділяють на одноосьові та багатоосьові, або фонтанні (див. рис. 23.6).
Всі нитчасті та гетеротрихальні Rhodophyta ведуть прикріплений спосіб життя. Прикріплення до субстрату здійснюється сланкими нитками, первинними ризоїдами та підошвами.
Розмноження та життєві цикли
Червоні водорості розмножуються нестатевим та статевим шляхами. Нестатеве розмноження відбувається вегетативно (поділом клітини надвоє, фрагментацією багатоклітинних таломів та додатковими "пагонами", які можуть відростати від сланких ниток або підошви) та за допомогою спор.
В останньому випадку розмноження здійснюється переважно моноспорами та тетраспорами (зрідка зустрічаються біспори та поліспори). Спори червоних водоростей позбавлені оболонки і здатні до амебоїдного руху. При утворенні спор в спорангіях у Rhodophyta, як правило, відбувається редукційний поділ, і, таким чином, спори гаплоїдні. Спори виходять назовні через верхівкову пору спорангію. Моноспори утворюються поодиноко з поверхневих (корових) клітин внаслідок відокремлення частини протопласту клітини-спорангія косою перегородкою; цей тип спорогенезу характерний для всіх бангіофіцієвих водоростей та найпримітивніших флоридей. Тетраспори утворюються по чотири; в залежності від напрямку площини поділу виділяють тетраспорангії хрестоподібні, зональні та тетраедричні (рис. 23.7).

Рис. 23.7. Спорангії червоних водоростей: а - моноспорангій; б-г - тетраспорангіїї (б - хрестоподібний, в - зональний, г - тетраедричний).
Тетраспорангії закладаються або серед клітин корового шару, або на поверхні талому в оточенні коротких захисних ниток - парафіз, утворюючи подушкоподібні структури - нематеції, або у спеціальних напіввідкритих заглибленнях - концептакулах. Тетраспори завжди проростають у гаплоїдні гаметофіти. У деяких бангієфіцієвих водоростей відомі також вкриті оболонкою апланоспори.
Статевий процес у червоних водоростей виключно оогамний, є майже у всіх представників, за вийнятком кокоїдних бангієфіцієвих та деяких флоридей. В чоловічих статевих органах - сперматангіях - утворюється по одному безджгутиковому спемацію, в жіночіх статевих органах - карпогонах - по одній яйцеклітині.
Карпогон, як правило, складається з розширеної частини - черевця, та трубкоподібного виросту - трихогіни, який уловлює спермації. У найпростішому випадку карпогони утворюються з будь-якої клітини талому, у більш складніших випадках - з верхівкової клітини спеціальних карпогоніальних ниток. Клітину, від якої відгалужується карпогоніальна нитка, називають несучою (ініціальною). У багатьох червоних водоростей з класу флоридей ця клітина надалі трансформується у спеціальну ауксилярну клітину, з якою карпогон зливається після запліднення.
При статевому процесі спермації виходять з сперматангіїв, пасивно переносяться током води, і у випадку потрапляння на карпогон запліднюють яйцеклітину. Одразу після запліднення зигота без періоду спокою проростає в особливе покоління, присутнє лише у Rhodophyta - карпоспорофіт. На карпоспорофіті утворюються карпоспори, які дають далі початок новому спорофіту (рис. 23.8).
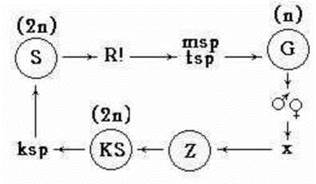
Рис. 23.8. Схема життєвого циклу Rhodophyta. S - спорофіт, G - гаметофіт, KS - карпоспорофіт, Z - зигота, msp - моноспори, tsp - тетраспори, ksp - карпоспори, gm - гамети, (2n) та (n) - диплоїдні та гаплоїдні набори хромосом, х - копуляція гамет.
Карпоспорофіт виконує особливу, спеціфічну функцію - він тиражує наслідки вдалого статевого процесу. Слід мати на увазі, що через відсутність джгутикових стадій ймовірність запліднення яйцеклітини чоловічою гаметою, що пасивно переноситься водою, є дуже низькою. Відповідно, і кількість запліднених яйцеклітин, що можуть дати початок новій рекомбінантній генерації, дуже мала, причому рівень смертності на початкових стадіях онтогенезу завжди підвищений. Забезпечення певного рівня рекомбінантної мінливості, достатнього для стабільного існування популяції, є функцією карпоспорофіту. Карпоспорофіт розвивається з заплідненого карпогону, і живиться за рахунок асимілятів, синтезованих гаметофітом. Клітини карпоспорофіту містять диплоїдні зиготичні ядра, що мітотично діляться. Врешті решт з клітин карпоспорофіту утворюються численні диплоїдні карпоспори, що надалі проростають у спорофіти.
Утворення карпоспор може відбуватися трьома основними шляхами, які обумовлюють три основні плани будови карпоспорофіту:
У найпростішому випадку зигота одразу починає ділитися, утворюючи карпоспори безпосередньо у карпогоні. В цьому варіанті карпоспорофіт дуже простий, і представлений карпогоном з карпоспорами.
При складнішому варіанті від карпогону відростають диплоїдні нитки з зиготичними ядрами - гонімобласти. Далі клітини таких гонімобластів перетворюються на карпоспори. Тут покоління карпоспорофіту представляють карпогон, гонімобласти та карпоспори.
Третій варіант найскладніший: карпогон з заплідненою яйцеклітиною зливається з ауксилярною клітиною, і утворює гетерокаріонтичну (різноядерну) клітину злиття. Остання містить одно або кілька власних гаплоїдних "гаметофітних" ядер, та диплоїдне зиготичне ядро, що потрапило у клітину злиття з заплідненого карпогону. Диплоїдне ядро кілька разів мітотично діляться, і дочірні ядра мігрують у периферичні випини клітини злиття. Кожний випин відокремлюється перегородкою, і проростає в гонімобласт, клітини якого перетворюються на карпоспори. В цьому варіанті карпоспорофіт складається з карпогону, клітини злиття, ауксилярних клітин, гонімобластів та карпоспор.
Таким чином, життєвий цикл у Rhodophyta переважно гаплодиплофазний, зі споричною редукцією і чергуванням трьох поколінь - гаметофіту, карпоспорофіту та спорофіту.
Система Rhodophyta
За будовою карпогону, хлоропластів, способами нестатевого розмноження, продуктами асиміляції, наявністю вторинних пор відділ поділяють на два класи - Bangiophyceae та Florideophyceae. Клас Bangiophyceae об'єднує водорості, у яких карпогон позбавлений трихогіни, карпоспори утворюються безпосередньо з зиготи, хлоропласти зірчасті або стрічкоподібні, як правило - з піреноїдом; продуктом асиміляції, крім багрянкового крохмалю, може бути також глікоген; нестатеве розмноження відбувається переважно моноспорами; первинні пори або позбавлені пробок, або мають прості пробки першого чи другого типу; вторинні пори відсутні. Більш продвинутий в еволюційному відношенні клас Florideophyceae включає водорості, у яких карпогон має трихогіну, карпоспори утворюються різними шляхами у різних порядках, хлоропласти численні, дископодібні, без піреноїдів; продуктом асиміляції є багрянковий крохмаль (глікоген не утворюється); первинні пори закриті пробками третього-сьомого типів (у порядку Ahnfeltiales пробкі прості, першого типу), клітини пов'язані між собою за допомогою як первинних, так і вторинних пор.
Клас Bangiophyceae
За типами морфологічної будови та розмноженням клас поділяють на 3 порядки - Porphyridiales, Compsogonales та Bangiales (табл. 23.1). Розподіл бангієфіцієвих водоростей по цим порядкам підтверджується також і молекулярними дендритами, побудованими на основі результатів секвенування генів rbcL та SSU rDNA, що кодують відповідно велику субодиницю RuBisCo та малу субодиницю цитоплазматичної рибосомальної РНК.
Таблиця 23.1. Деякі фенотипічні ознаки представників порядків Bangiophyceae
Порядок |
Тип структури |
Статевий процес |
Таломи |
Поширення |
Porphyridiales |
кокоїдний |
відсутній |
різноманітні слизові колонії |
ґрунти, прісні водойми |
Compsogonales |
гетеротрихальний |
відсутній |
ниткоподібні розгалужені |
прісні водойми |
Bangiales |
нитчастий |
наявний |
у формі пластин або багаторядної нитки |
моря |
Porphyridiales об'єднує кокоїдні водорості, які, як правило, утворюють слизові колонії. Розмножуються поділом клітин надвоє, моноспорами або апланоспорами. Останні утворюються в клітині-спорангії по чотири, але на відміну від тетраспор одягнуті клітинною оболонкою. Спеціальні цитологічні дослідження одного з представників цього порядку (Cyanidium) показали, що цитокінез здійснюється шляхом утворення кільцевої борозни, причому ріст цієї борозни обумовлений скороченням периферичного білкового кільця, побудованого з скоротливого білку актину.
Порфіридіальні водорості мешкають переважно у прісних водоймах та у ґрунті. Найбільш поширеним є рід Porphyridium (рис. 23.9). Види цього роду інтенсивно досліджується у багатьох лабораторіях, оскільки вважаються перспективними фікотехнологічними об'єктами для промислового культивування з метою отримання фікоколоїдів.
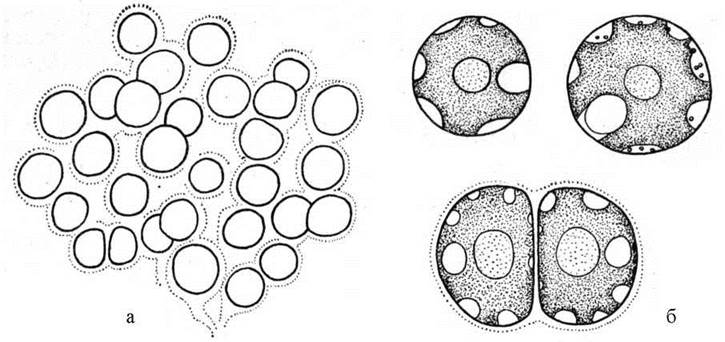
Рис. 23.9. Слизова колонія (а) та окремі клітини (б) Porphyridium purpureum.
Таломи представників порядку Compsopogonales гетеротрихальні, і завичай мають вигляд розгалужених кущиків. Розмножуються моноспорами, статевий процес невідомий. Компсогональні водорості мешкають переважно у прісних водоймах тропічного поясу. На Україну ці водорості завезені разом з тропічними акваріумними рослинами. Зокрема, в акваріумах часто зустрічається Compsopogon, відомий в акваріумістів під назвою "синя борода". Водорість розвивається як епіфіт, переважно на рослинах-криптокорінах (рис. 23.10).
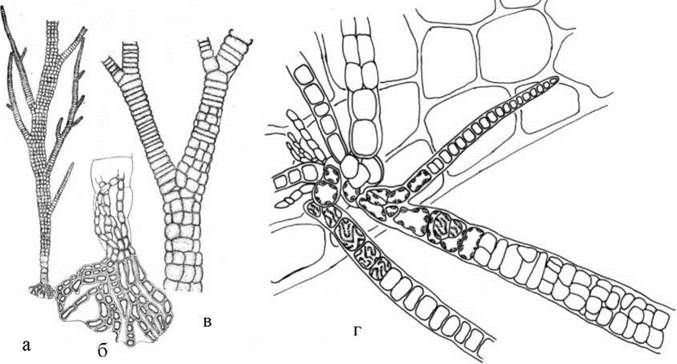
Рис. 23.10. Compsopogon: а - зовнішній вигляд талому; б - базальна частина талому з ризоїдальними нитками, зануреними у тканини рослини-криптокоріни; в - висхідна частина нитки; г - фрагмент талому (показані клітини, що містять стрічкоподібні хлоропласти, у багаторядній висхідній частині спостерігаються поодинокі моноспорангії, що мають трикутно-заокруглену форму).
Bangiales. До цього порядку входять водорості, які мають нитчасті, як правило - багаторядні нерозгалужені або пластинчасті таломи. Розмножуються бангіальні моноспорами (у деяких видів - поліспорами) та додатковими "пагонами". Статевий процес відомий, і характеризується зиготичною редукцією, яка відбувається одразу ж після запліднення яйцеклітини. Представники порядку живуть переважно у морях. Найбільш поширеними є роди Bangia та Porphyra. Bangia має багаторядний нитчастий талом. У Porphyra талом пластинчастий. Porphyra (т.з. "червоний морський салат") є їстівною водорістю, і введена у промислову аквакультуру (зокрема вирощується у Чорному морі) (рис. 23.11).
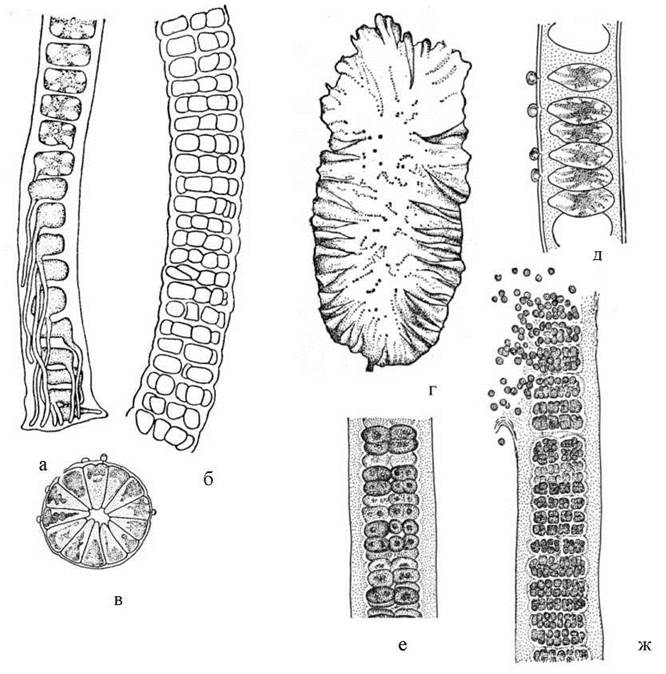
Рис. 23.11. Бангіальні водорості: а-в - Bangia (а - базальна частина талому з ризоїдами, б - середня багаторядна частина талому, в - поперечний зріз через талом в зоні розвитку моноспорангіїв); г-ж - Porphyra (г - зовнішній вигляд талому, д-ж - поперечний зріз через пластину в зоні карпогонів (д), сперматангіїв (е) та карпоспорангіїв (ж) (за Виноградова, 1977).
Клас Florideophyceae
Провідний клас у системі Rhodophyta, який об'єднує біля 80% відомих видів відділу. До класу входять водорості виключно з гетеротрихальним типом будови. За молекулярними даними, в межах класу представлено п'ять досить відмінних на молекулярному рівні груп, які можуть відповідати таксонам рангу порядків або груп порядків. З молекулярною уособленістю кожної групи добре корелює комплекс фенотипічних ознак, пов'язаних з типами пробок первинних пор, наявністю та особливостями ауксилярних клітин, наявністю карпогоніальних гілок, типом чергування поколінь, морфологією тетраспор, екологічними особливостями. Різні автори виділяють у межах класу від шести до чотирнадцяти порядків, з яких нижче розглядаються вісім: Hildenbrandiales, Nemaliales, Corallinales, Ahnfeltiales, Gelidiales, Gigartinales, Rhodymeniales, Ceramiales.
Hildenbrandiales вважається найпримітивнішим порядком флоридей. Представники порядку мешкають як у морях, так і в прісних водоймах. Таломи цих водоростей розвиваються переважно на кам'янистому субстраті і утворюють на його поверхні шкуринки псевдопаренхіматозної будови, що складаються з кількох шарів клітин. Первинні пори закриті пробками третього типу. Ці пробки не мають поверхневої мембрани, над білковою основою є два додаткових полісахаридних шари, причому зовнішній шар дуже товстий та опуклий. Статевий процес відсутній, життєвий цикл являє собою цикломорфоз. Розмноження лише вегетативне: фрагментацією таломів, групами вегетативних клітин (геммами) та сланкими нитками - столонами. Типовим представником є рід Hildenbrandia. Вважається, що від Hildenbrandiales беруть початок дві різні екологічні групи флоридей. Перша гілка - порядок Nemaliales - включає водорості, в межах яких спостерігається перехід від морського до прісноводного способу існування. Другу гілку представляють майже виключно морські види всіх інших порядків даного класу.
Для Nemaliales характерними ознаками є наявність пробок шостого або сьомого типів: з поверхневою мембраною та двома полісахаридними шарами, причому зовнішній шар може бути як тонким, так і товстим. Таломи у немаліальних переважно гетеротрихальні або псевдопаренхіматозні, як одно-, так і багатоосьові. Нестатеве розмноження здійснюється тетраспорами, що утворюються в хрестоподібних тетраспорангіях. Ауксилярні клітини відсутні. Карпогони можуть утворюватися як на коротких карпогоніальних гілочках, так і зі звичайних вегетативних клітин. Життєві цикли гаплодиплофазні, зі споричною редукцією та зміною поколінь, ізоморфних або гетероморфних. Характерними представниками є роди Palmaria, Nemalion та Batrachospermum.
Palmaria мешкає виключно у морях. Спорофіти та чоловічі гаметофіти у цієї водорості морфологічно однакові, мають вигляд розсічених пластин псевдопаренхіматозної багатоосьової будови. Жіночі гаметофіти дрібні, коркоподібні, складаються з кількох шарів клітин. Карпогони утворюються з клітин поверхневого шару. Після запліднення яйцеклітина одразу проросте в спорофіт, не утворюючи гонімобластів. Спорофіт при цьому залишається прикріпленим до жіночого гаметофіту. На зрілому спорофіті розвиваються хрестоподібні тетраспорангії з гаплоїдними тетраспорами, які надалі дають початок чоловічім або жіночім гаметофітам. Таким чином, у Palmaria життєвий цикл гаплодиплофазний, зі споричним мейозом, гетероморфною зміною поколінь, статевим диморфізмом та без стадії карпоспорофіту. Досить поширений представник - Palmaria palmata - є їстівною морською водорістю, що належить до так званих червоних морських салатів.
Більшість видів роду Nemalion також мешкає у морях, проте окремі представники зустрічаються у солонуватих та навіть прісних водоймах. Гаметофіти Nemalion макроскопічні, мають вигляд невеликих кущиков. Таломи гаметофітів багатоосьові, складаються з групи опірних осьових ниток, від яких відгалужуються численні бічні нитки-асимілятори. З верхівкових клітин асиміляторних ниток розвиваються або численні одноклітинні антеридії - сперматангії, з яких утворюються спермації, або трьох-п'ятиклітинні карпогоніальні гілочки, на верхівці яких закладаються карпогони із трихогінами. Після запліднення зигота без періоду спокою ділиться, і утворює щільну групу малоклітинних гонімобластів, у яких верхівкові клітини перетворюються на карпоспори.
Карпоспори проростають в одноосьові розгалужені кущисті спорофіти. Восени та взимку у верхівкових клітинах бічних гілочок відбувається мейоз, і з них утворюються хрестоподібні тетраспорангії. Тетраспори далі проростають у нові багатоосьові гаметофіти.
У Batrachospermum гаплоїдний макроскопічний гаметофіт розвивається на диплоїдному мікроскопічному спорофіті (рис. 23.12). Спорофіт у цієї водорості представлений однорядними розгалуженими нитками, у верхівкових клітинах яких може відбуватися мейоз. Після мейозу такі клітини не перетворюються на тетраспорангії, а починають безпосередньо проростати у спорофіти. Таломи спорофітів оточені товстим шаром слизу, диференційовані на однорядну опірну нитку та мутовчасто розміщені бічні гілочки-асимілятори. Сперматангії утворюються з апікальних клітин асиміляторних ниток, карпогони - на коротеньких карпогоніальних гілочках, які у різних видів розміщуються на різних частинах талому гаметофіту. Після запліднення з карпогону розвивається карпоспорофіт, схожий з карпоспорофітом Nemalion. Всі види роду Batrachospermum є прісноводними водоростями, що мешкають переважно у холодних гірських річках або в великих озерах. Таломи цих водоростей можуть мати різні відтінки як червоного, так і синьо-зеленого забарвлення.

Рис. 23.12. Batrachospermum: а - диплоїдний спорофіт; на верхівці розташовуються клітини-спорангії та тетради клітин, гомологічні тетраспорангіям, з яких починається розвиток гаметофіту; б - зовнішній вигляд гаметофіту; в - фрагмент гаметофіту зі сферичними карпоспорофітами.
У системах ряду авторів Palmaria, Nemalion та Batrachospermum розміщуються у окремих самостійних порядках (Palmariales, Nemaliales та Batrachospermatales, відповідно) через відміни у життєвих циклах, деталях розмноження, будові таломів та екології. Проте молекулярні та цитологічні дослідження свідчать про тісні родинні зв'язки цих представників, незважаючи на їх відмінність на морфологічному рівні.
Найяскравішою особливістю Corallinales є кальцифікація клітинних покривів. Таломи багатоосьові, звичайно просякнуті вапном і мають вигляд червоного каміння, невеликих кущиків або нагадують корали. Пробки первинних пор належать до третього типу - вони позбавлені поверхневої мембрани, мають два додаткових шара полісахаридів, причому зовнішній шар дуже товстий. Чергування поколінь у представників порядку ізоморфне. Органи як нестатевого, так і статевого розмноження (тетраспорангії, карпогони та сперматангії) розвиваються у спеціальних заглибинах талому, що називаються концептакулами. Тетраспорангії хрестоподібні або зональні.
Карпогон закладається на верхівці двоклітинної карпогоніальної нитки. Клітина, розташована під карпогоном, називається ообластемною. Карпогоніальна нитка, утворена карпогоном та ообластемною клітиною, бере початок від несучої (ініціальної) клітини, від якої також розвивається ще одна коротка, проте стерильна нитка. Ініціальна клітина надалі перетворюється на ауксилярну клітину (рис. 23.13).
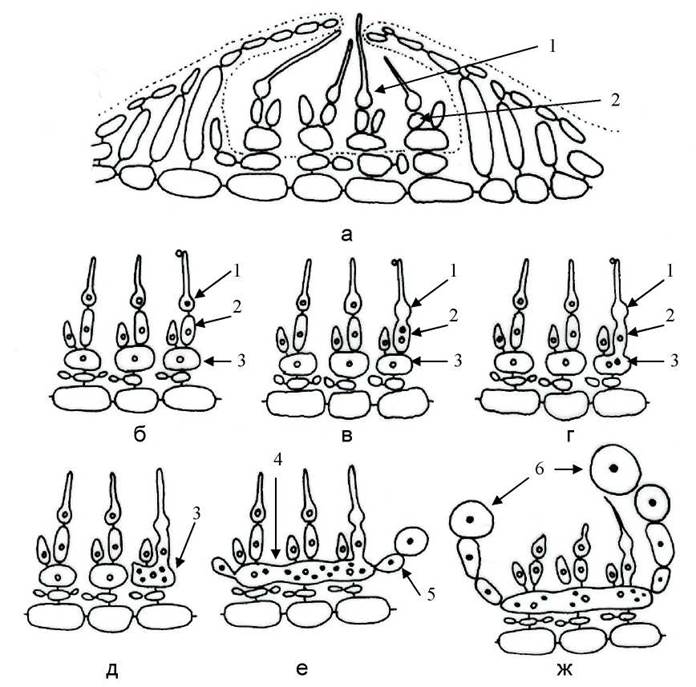
Рис. 23.13. Будова концептакулу з карпогонами (а) та послідовні стадії розвитку карпоспорофіту представників Corallinales (б-ж): 1 - карпогон з трихогіною; 2 - ообластемна клітина; 3 - ауксилярна клітина; 4 - гетерокаріонтична клітина злиття, що утворилась внаслідок плазмагамії ауксилярних клітин; 5 - гонімобласт; 6 - карпоспора. Диплоїдні зиготичні ядра чорні, гаплоїдні ядра вегетативних клітин всередині світлі.
Після запліднення карпогон зливається з ообластемною клітиною, яка, в свою чергу, зливається з клітиною ауксилярною. Через ообластемну клітину диплоїдне ядро зиготи мігрує в ауксилярну клітину. Далі всі ауксилярні клітини одного концептакулу зливаються між собою, і в концептакулі утворюється одна велика багатоядерна гетерокаріонтична клітина злиття, яка містить диплоїдні зиготичні ядра та гаплоїдні вегетативні ядра. Диплоїдні ядра мігрують у бічні випини клітини злиття, і відокремлюються від останньої клітинними перегородками. Далі такі диплоїдні клітини діляться, перетворюючись на гонімобласти, з верхівкових клітин яких розваваються карпоспори. Зрілі карпоспори через отвір виходять з концептакулу назовні, і проростають у спорофіти.
Найпоширенішими родами порядку є Lithothamnion та Corallina. У Lithothamnion таломи коркоподібні або мають вигляд більш-менш розгалуженого червоного вапнякового каміння (рис. 23.14).
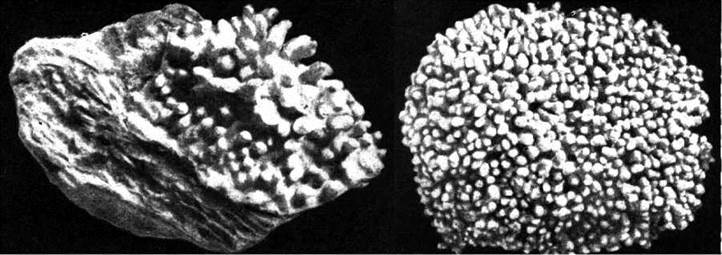
Рис. 23.14. Зовнішній вигляд таломів Lithothamnion.
Для Corallina характерні таломами у вигляді пірчасто та вильчасто розгалужених кущиків. Кожна гілка талому складається з вапнякових сегментів-члеників, які утворені багаторядними опірними нитками та бічними нитками-асиміляторами (рис. 23.15). Ділянки між сусідніми сегментами позбавлені асиміляторів, а клітини опірних ниток досить видовжені. Обидва роди зустрічаються у різних морях, зокрема в Чорному морі.
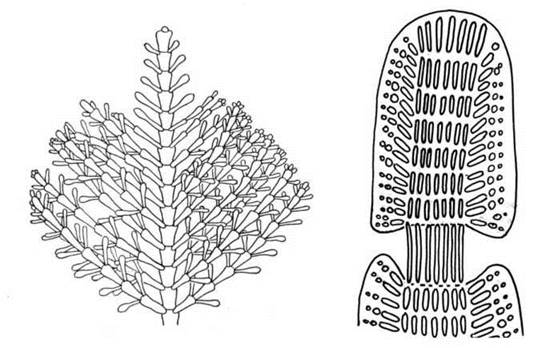
Рис. 23.15. Corallina. Фрагмент талому (ліворуч) та ділянки, що з'єднує два членики-сегменти (праворуч).
До порядку Ahnfeltiales належить лише один рід - Ahnfeltia, або морський мошок (рис. 23.16). У анфельції таломи не просякуються вапном, пробки первинних пор належать до першого типу - вони позбавлені як поверхневої мембрани, так і полісахаридних шарів. Таломи мають вигляд дихотомічно розгалужених кущиків псевдопаренхіматозної будови. Статевий процес не виявлений, і життєвий цикл представлений цикломорфозом. Розмноження, на відміну від інших флоридей, здійснюється моноспорами, утворенню яких не передує мейоз. Моноспори розвиваються у нематеціях - поверхневих потовщеннях талому, утворених короткими вертикальними нитками, що відходять від корових клітин. Ahnfeltia масово зустрічається у північних морях, і є основним джерелом для отримання біломорського агар-агару.
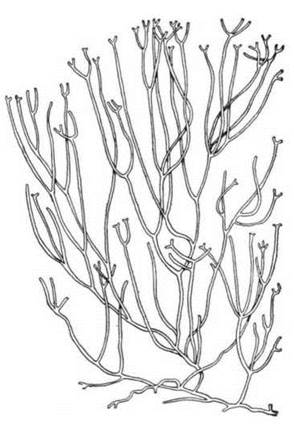
Рис. 23.16. Зовнішній вигляд талому Ahnfeltia.
У чотирьох наступних порядків - Gelidiales, Gigartinales, Rhodymeniales та Ceramiales - є багато спільних рис: пробки первинних пор мають поверхневу мембрану, і або взагалі позбавлені полісахаридних шарів, або мають лише один тонкий полісахаридний шар (четвертий та п'ятий типи пробок), карпогони розвиваються з верхівкової клітини триклітинних карпогоніальних ниток; запліднені карпогони зливаються безпосередньо з ауксилярною клітиною, без участі ообластемних клітин. Зміна поколінь ізоморфна, і лише у деяких представників Gigartinales - гетероморфна. Відміни між цима еволюційно близькими порядками полягають у планах будови таломів, типах тетраспорангіїв, наявності полісахаридного шару у пробці, а також пов'язані з особливостями розвитку ауксилярних клітин (зокрема, з місцем їх формування та з часом: до чи після запліднення карпогону).
Для представників Gelidiales характерні одноосьові таломи псевдопаренхіматозної будови, у яких пори між клітинами закриті пробками п'ятого типу; тетраспорангії хрестоподібні, ауксилярні клітини утворюються з ініціальних клітин, до запліднення карпогону. Карпогоніальні гілочки розміщуються в глибині таломів, і за межі назовні виходять лише трихогіни карпогонів. Після запліднення карпогон зливається з ауксилярною клітиною, утворюючи гетерокаріонтичну клітину злиття, в якій диплоїдне ядро кілька разів мітотично ділиться (рис. 23.17). Далі у бічні вирости клітини злиття заходять диплоїдні ядра, вирости відокремлюються перегородками, і розвиваються у гонімобласти. На цій стадії навколо карпоспорофіту суцільним шаром розростаються численні дрібні трофічні клітини, і, як наслідок, утворюється цистокарпій, що має вигляд здуття або невеличкої сферичної пухлини на таломі гаметофіту (рис. 23.18).

Рис. 23.17. Послідовні стадії розвитку карпоспорофіту представників Gelidiales: а - запліднення; б - утворення клітини злиття; в - утворення гонімобластів та торфічних клітин; г - цистокарпій. 1 - карпогон з трихогіною; 2 - ауксилярна клітина; 3 - базальна клітина карпогоніальної нитки; 4 - гетерокаріонтична клітина злиття; 5 - початкова стадія розвитку гонімобласту; 6 - трофічні клітини; 7 - зрілі клітини гонімобластів; 8 - карпоспора; 9 - шар трофічних клітин цистокарпію. Диплоїдні зиготичні ядра чорні, гаплоїдні ядра вегетативних клітин всередині світлі.

Рис. 23.18. Gelidium: a - зовнішній вигляд талому; б - фрагмент талому з цистокарпіями; в, г - зрізи через апікальну (в) та медіальну (г) частини талому.
Типовий представник порядку - Gelidium latifolium - має досить великі (до 10 см заввишки) сплощені пірчасті псевдопаренхіматозні таломи одноосьової будови, які часто можна знайти на камінні у літоральній зоні морів помірного поясу (в тому числі, в Чорному морі).
У водоростей з порядку Gigartinales таломи також псевдопаренхіматозні, одно- або багатоосьові; пробки належать до четвертого типу, тетраспорангії хрестоподібні або зональні, розвиваються переважно в нематеціях. Карпогон подібно до попереднього порядку, розвивається з верхівкової клітини карпогоніальної нитки, ауксилярна клітина - з ініціальної клітини. Після запліднення карпогон зливається з ауксилярною клітиною, утворюючи гетерокаріонтичну клітину злиття. З бічних виростів цієї клітини формуються гонімобласти, а з них у свою чергу - карпоспори (рис. 23.19). Гігартинальні водорості цікаві, в першу чергу, тим, що до них належить більшість видів, з яких у промислових масштабах отримують фікоколоїди.
Зокрема, з видів роду Phyllophora (рис. 23.20) отримують агароїд, який називають "чорноморським агаром". Уздовж північно-західного узбережжя Чорного моря, між Одесою та Очаковим знаходиться зона, де на глибині 5-60 м суцільні зарості утворює Phyllophora nervosa. Це найбільший у світі філофорний фітоценоз, який називають філофорним полем Зернова, на честь його першовідкривача - видатного морського гідробіолога академіка *.*. Зернова. Завдяки філофорному полю Зернова Україна є державою з найбільшими у світі запасами промислової сировини для отримання агароїду.
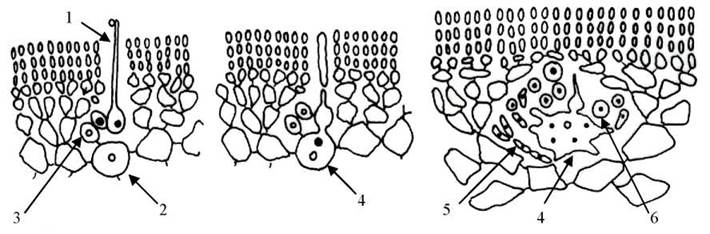
Рис. 23.19. Послідовні стадії розвитку карпоспорофіту представників Gigartinales (на прикладі Chondrus). 1 - карпогон з трихогіною; 2 - ініціальна клітина карпогоніальної гілки; 3 - вегетативна клітина карпогоніальної гілки; 4 - гетерокаріонтична клітина злиття; 5 - гонімобласт; 6 - карпоспора. Диплоїдні зиготичні ядра чорні, гаплоїдні ядра вегетативних клітин всередині світлі.
Таломи Phyllophora кущисті, представлені сланкими "пагонами", від яких підіймаються вертикальні стебельця. Верхня частина кожного стебельця сплощена, розростається по краям, і утворює пластину з потовщеною середньою частиною - ребром. Пластина рясно галузиться, і часто по краям та на ребрі несе дочірні, молодші пластини. Тетраспорангії та статеві органи розвиваються у нематеціях, розташованих на поверхні пластин вздовж ребра.

Рис. 23.20. Phyllophora nervosa, що є основним джерелом отримання агароїду в Україні.
Основним джерелом сировини для отримання карагеніну є дві досить схожі водорості, що мешкають у північній частині Атлантичного океану та у морях Північного льодовитого океану - Chondrus, або ірландський мох, та Gigartina. Обидві водорості мають сплощені, рясно дихотомічно розгалужені таломи (рис. 23.21). Проте у Chondrus поверхня таломів гладенька, тоді як у Gigartina вона вкрита додатковими численними невеличкими виростами.

Рис. 23.21. Гігартинальні водорості: а-в - Chondrus (а - зовнішній вигляд талому, б - зріз через спорофіт з тетрасорангіями; в - гаметофіт з цистокарпіями); г - зовнішній вигляд талому Gigartina.
Псевдопаренхіматозні таломи, переважно пластинчастої форми та з порожниною всередині, характерні для представників Rhodymeniales. Будова таломів виключно багатоосьова, пори між клітинами закриті пробками четвертого типу, тетраспорангії хрестоподібні та тетраедричні. Ініціальна клітина, з якої починається розвиток жіночих статевих органів, дає початок двом ниткам - триклітинній карпогоніальній, та двоклітинній ауксилярній, з ауксилярною клітиною на верхівці (рис. 23.22). Система з ініціальної клітини та карпогоніальної й ауксилярної ниток отримала назву прокарпія. Вегетативні клітини, що знаходяться поруч із прокарпієм, під час розвитку гонімобластів, розростаються, і утворюють асиміляційний та трофічний покрив цистокарпію. У процесі дозрівання карпоспор цистокаріпії збільшуються у розмірі, і на таломах гаметофітів утворюють сферичні здуття.
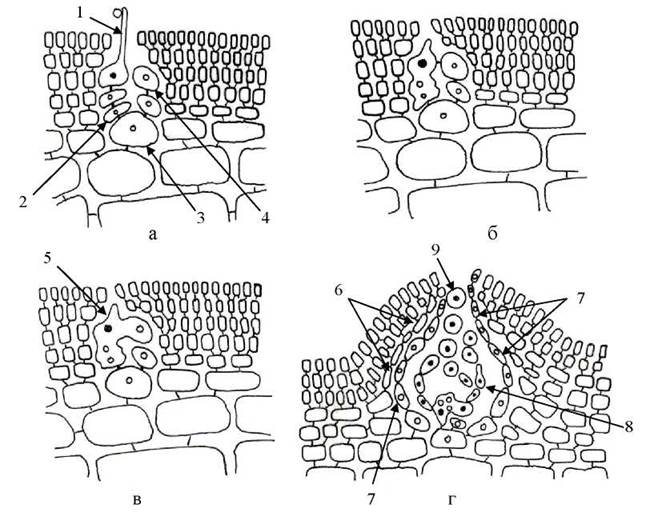
Рис. 23.22. Послідовні стадії розвитку карпоспорофіту Rhodymenia: а - прокарпій; б - плазмагамія між клітинами карпогоніальної гілки; в - утворення клітини злиття; г - цистокарпій. 1 - карпогон з трихогіною; 2 - базальна клітина карпогоніальної гілки; 3 - ініціальна клітина; 4 - ауксилярна клітина; 5 - гетерокаріонтична клітина злиття; 6 - клітини асиміляторного шару
цистокарпію; 7 - клітини торфічного шару цистокарпію; 8 - гонімобласт; 9 - карпоспора. Диплоїдні зиготичні ядра чорні, гаплоїдні ядра вегетативних клітин всередині світлі.
Типовий представник порядку - Rhodymenia - поширений в літоральній зоні північних морів (рис. 23.23). Родіменія, як і порфіра, є їстівною багрянкою, і навіть має ту ж саму народну назву - червоний морський салат.

Рис. 23.23. Зовнішній вигляд таломів Rhodymenia stenogona (ліворуч) та R. pertusa (праворуч). (За Виноградова, 1977).
Водорості порядку Ceramiales мають ниткоподібні одноосьові таломи, що прикріплюються до субстрату підошвою, яку утворює базальна клітина; сланкі нитки таломів редуковані. Цераміальним притаманні пробки четвертого типу, тетраедричні тетраспорангії, прокарпії та цистокарпії схожої з Rhodymeniales будови. Розвиток карпоспорофіту також нагадує попередній порядок, але ауксилярні клітини утворюються лише після запліднення яйцеклітини (рис. 23.24). Порядок є найчисленнішим в межах відділу, і за системами різних авторів нараховує 1000-1500 видів.
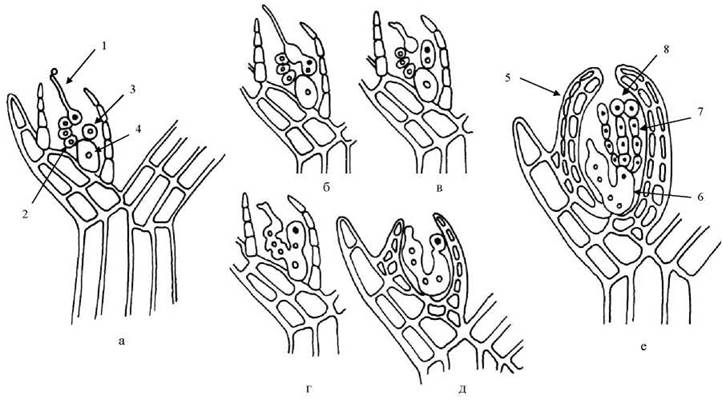
Рис. 23.24. Послідовні стадії розвитку карпоспорофіту Polysiphonia: а - прокарпій; б - г - послідовні стадії утворення гетерокаріонтичної клітини злиття; д - початкова стадія утворення гонімобластів та покриву цистокарпію; е - цистокарпій. 1 - карпогон з трихогіною; 2 - базальна клітина карпогоніальної гілки; 3 - ауксилярна клітина; 4 - ініціальна клітина; 5 - асиміляторний та трофічний шари цистокарпію; 6 - гетерокаріонтична клітина злиття; 7 - гонімобласти; 8 - карпоспора. Диплоїдні зиготичні ядра чорні, гаплоїдні ядра вегетативних клітин всередині світлі.
Характерними родами водоростевих угрупувань літоралі та субліторалі різних морів є роди Callithamnion, Ceramium, Polysiphonia, Odonthalia.
Callithamnion має кущисті таломи, утворені однорядними рясно розгалуженими нитками. Тетраспорангії, сперматангії та карпогони утворюються в пазухах верхівкових гілочок; цистокарпії переважно парні (рис. 23.25). Представники роду характерні для морів помірного поясу, масово зустрічаються у Чорному та Азовському морях, де розвиваються переважно у бентосних та перифітонних угрупуваннях на глибинах до 3 -5 м.
Види роду Ceramium також мають вигляд невеличких кущиків. Нитки галузяться дихотомічно, верхівки вильчасті, і загнуті всередину. Талом утворений однорядною опірною ниткою, яка у зоні контакту суміжних клітин вкрита корою. У деяких видів кора вкриває весь талом, проте частіше медіальна частина клітин опірних ниток лишається відкритою, через що рослина набуває членистого вигляду - прозорі відкриті ділянки опірних клітин чергуються з ділянками, вкритими корою, що забарвлена в інтенсивний червоний колір. Кора являє собою асиміляторні короткі нитки, що щільно притискаються до оболонок клітин опірних ниток. Деякі асиміляторні нитки видозмінюються у щетинки або шипи. З клітин кори також розвиваються сперматангії, карпогоніальні нитки та тетраспорангії (рис. 23.25).
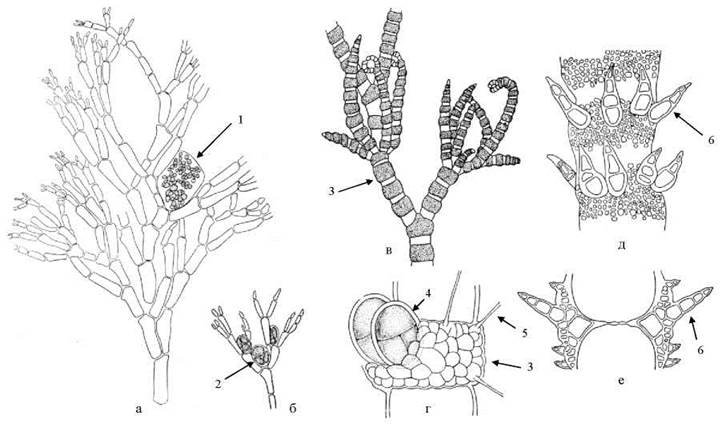
Рис. 23.25. Цераміальні водорості: Callithamnion (а, б) та Ceramium (в-е). Фрагмент талому з цистокарпієм (а); верхівкова гілка з тетраспорангіями (б); фрагмент талому з поясками (в); поясок з тетраспорангіями та щетинками (г); зона пояску з шипами (д, е). 1 - цистокарпій; 2, 4 - тетраспорангії; 3 - поясок; 5 - щетинка; 6 - шип.
Ceramium зустрічається в усіх широтах, але масово розвивається переважно у теплих морях, зокрема в Чорному та Азовському.
Ще одним дуже поширеним представником порядку є Polysiphonia. Таломи цієї водорості також кущисті, і мають членисту будову, але іншого типу, ніж у Ceramium. Нитку утворюють два типи клітин - центральні, що виконують опірну функцію, та перицентральні, які здійснюють фотосинтез і утворюють репродуктивні органи - тетраспорангії, сперматангії та карпогони. Перицентральні клітини розташовуються правильними ярусами, причому один ярус перицентральних клітин оточує однорядним кільцем одну центральну клітину. Перицентральні та центральну клітини одного ярусу називають, відповідно, периферичними та центральним сифонами.
Таломи полісифонної будови характерні також для роду Odonthalia, що поширений у холодних північних морях. Проте на відміну від полісіфонії, дві групи периферичних сифонів, розташованих один навпроти одного з двох боків центрального сифону, інтенсивно діляться в поздовжному напрямку. Як наслідок, протилежні боки нитки розростаються, і поступово набувають пластинчастої форми. Інколи талом навколо центральної частини потовщується, утворюючи центральний нерв.
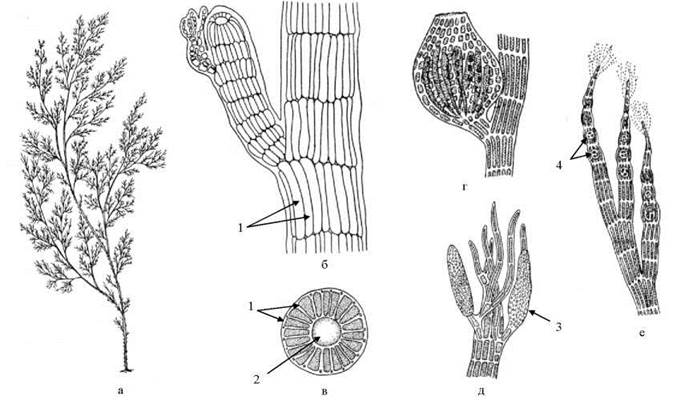
Рис. 23.26. Polysiphonia: а - загальний вигляд талому; б - фрагмент талому з короткими бічними гілочками; в - поперечний зріз талому; г - цистокарпій; д - гілка зі сперматангіями; е - гілка з тетраспорангіями. 1 - перицентральні клітини (сифони); 2 - центральна клітина; 3 - сперматангій; 4 - тетраспорангії.
Напрямки еволюції відділу
Еволюція червоних водоростей пов'язана, перш за все, з еволюцією карпоспорофіту. Оскільки статевий процес відбувається шляхом запліднення яйцеклітини пасивним, нерухомим, не здатним до таксису спермацієм, ефективність його була б невисокою внаслідок обмеженої кількості вдалих актів запліднення при значних витратах ресурсів на збільшення ймовірності таких актів (збільшення кількості карпогонів та сперматангіїв). У флоридей ця проблема вирішується оригінальним шляхом - ефективність статевого процесу підвищується за рахунок збільшення продукції карпоспор у кожному вдалому статевому акті. У примітивних форм (Bangiales) зигота безпосередньо утворює карпоспори, але продукційні можливості зиготи обмежені її розмірами (зокрема, при великій кількості дочірніх ядер буде порушуватися плазмено-ядерне відношення у карпоспорах). Проблему плазмено-ядерного співвідношення розв'язує поява гонімобластів, які утворюють карпоспори (Nemaliales).
Проте обмеження тепер ставить нестача поживних речовин у зиготі, з якої гонімобласти розвиваються. Наступним кроком вперед є забезпечення гонімобластів поживними речовинами, що накопичують спеціалізовані трофічні системи - ауксилярні клітини. У більш примітивніших порядків (Corallinales) зиготичне ядро потрапляє в ауксилярну клітину через ообластемну клітину, після її злиття з карпогоном та ауксилярною клітиною. У більш продвинутих в еволюційному відношенні порядків карпогон зливається з ауксилярною клітиною поминаючи клітини-"посередники" (Gelidiales, Gigartinales, Rhodymeniales).
Додаткове живлення гонімобластів також забезпечують оточуючі карпоспорофіти вегетативні клітини, що видозмінюються у трофічні клітини оболонок цистокарпіїв. Найбільша ефективність використання ресурсу поживних речовин для розвитку карпоспорофіту досягається тоді, коли ауксилярна клітина утворюється лише після запліднення (Ceramiales). В цьому випадку поживні речовини не залишаються законсервованими в ауксилярних клітинах біля незапліднених карпогонів, а направляються лише туди, де відбувся вдалий статевий акт.
Поширення
Червоні водорості поширені переважно у бентосних та перифітонних морських угрупуваннях. Найбільш різноманітні та численні Rhodophyta у тропіках та субтропіках, при наближенні до полюсів видове багатство червоних водоростей зменшується. У теплих морях представлені водорості переважно дрібних розмірів. У холодних морях більшість Rhodophyta має великі, макроскопічні таломи. Подібна тенденція простежується і в розподілі червоних водоростей за глибиною - представники з великими таломами зустрічаються на більших глибинах. Серед Rhodophyta багато глибоководних видів (нижня межа трапляння - 200 м). Вважають, що глибоководні представники використовують енергію проникливих коротких хвиль фіолетової та ультрафіолетової частин спектру і тому відбивають довгохвильові червоні промені.
Досить значна кількість видів бангіофіцієвих та деякі примітивні флоридеї мешкають у прісних водоймах (Compsopogon, Batrachospermum, Chantransia). На відміну від морських, прісноводні багрянки забарвлені у синій або сіро-сталевий колір. Деякі Porphyridiales мешкають у позаводних умовах, зокрема - у ґрунті (Porphyridium).
Чимало морських Rhodophyta паразитує на інших червоних водоростях. Цікаво, що біля 90% паразитичних червоних водоростей мають господарів, які у систематичному відношенні є близько спорідненими з паразитом. Такий тип паразитизму називають адельфопаразитизмом.
При цьому типі відношень паразит починає розвиватися на поверхні господаря як епіфіт, далі одна з клітин утворює відросток, що проникає в цитоплазму господаря. У зоні контакту оболонка відростку та господаря розчиняється, і ядро, мітохондрії та рибосоми паразита потрапляють в цитоплазму господаря. Ядро паразита ділиться швидше, ніж ядро господаря. Уражена клітина формує нові відростки, через які ядра, мітохондрії та рибосоми адельфопаразита потрапляють в інші клітини. Таким чином, невдовзі цитоплазма більшої частини клітин господаря заміщується цитоплазмою паразита.
Незначна кількість червоних водоростей паразитує на господарях, які еволюційно віддалені від паразиту, хоча теж належать до Rhodophyta. Цей тип паразитизму називають аллопаразитизмом.
Значення у природі
У морських фітоценозах червоні водорості, як правило, є субдомінантами і за біомасою та продукцією поступаються морським бурим водоростям. Проте у деяких випадках Rhodophyta є провідною групою продуцентів на досить значних акваторіях (наприклад, у районі філофорного поля Зернова).
Звичайно Rhodophyta складають харчову базу багатьох гідробіонтів. Для захисту від надмірного виїдання деякі червоні водорості (наприклад, Laurencia) продукують токсичні речовини. Тварини, які стійкі до дії цих токсинів (зокрема, морські зайці з роду Aplysia) концентрують ці токсини як засіб захисту від хижаків. У тропіках криптонеміальні водорості своїми вапняковими таломами"прошивають" гілки коралів і таким чином закріплюють коралові рифи. Види роду Lithothamnion у різних морях утворюють літотамнієві вапняки.
Практичне використання
У практичній діяльності людина використовує червоні водорості переважно у трьох напрямках - як джерело фікоколоїдів, у їжу та для отримання лікарських препаратів.
Фікоколоїди (агар, агароїд, карагенін) є найбільш цінними продуктами, які отримують з червоних водоростей. Фікоколоїди являють собою сульфатовану та піруватовану галактозу; 1-2% розчини фікоколоїдів при кімнатній температурі застигають, утворюючи тверді гелі. Джерелом агару для країн північної частини Евразії є Ahnfeltiaplicata, яка мешкає у північних морях. У країнах субтропічного та тропічного поясів агар отримують переважно з Gracilaria та Gelidium. Сировиною для виробництва агароїду є Phyllophora (в Україні з неї виробляють т.з. чорноморський агар). Найбільшим постачальником карагеніну на світовому ринку є Філіппіни, де в аквакультуру введено водорості з роду Euchemia. У північних країнах карагенін отримують переважно з Chondrus.
Агар використовують для виготовлення твердих поживних середовищ, як желюючий компонент у кондитерській промисловості, з агароїду отримують агарозу - основний гель для електрофорезу та хроматографії, карагенін застосовують як емульгатор у фармацевтиці та виробництві молочних продуктів, як клеючу речовину у текстильному та шкіряному виробництвах.
Харчовими водоростями є переважно червоні морські салати - Porphyra та Rhodymenia: першу вирощують на спеціалізованих морських фермах (в тому числі і в Чорному морі), другу збирають як супутній продукт при промислі морської капусти. Порфіра вважається делікатесом, смак якої обумовлюють вільні амінокислоти та ізофлоризиди.
Основними фармацевтичними препаратами, які виготовляють з червоних водоростей, є препарати для лікування підвищеної кислотності (отримують з криптонеміальних водоростей, переважно - з Corallina officinalis та Lithothamnion), глистогонні засоби (Digenia), гемагглютиніни, специфічні до групи В крові людини (Ptilota plumosa).
Крім того, червоні водорості використовують як органічні добрива, як біоіндикатори для оцінки якості води.