Альгологія - Костіков І.Ю. - 2009-2013
Розділ 24. Зелені водорості – Chlorophyta
Відділ зелених водоростей (Chlorophyta або Chlorophycophyta) є найчисленішим серед водоростей. За даними різних авторів, він нараховує 20-25 тис. видів. Найдавніші рештки відомі з верхнього докембрію - протерозою (біля 1400 млн.років). Зелені водорості поширені в прісних та гіпергалінних водоймах, в морях та океанах, в наземних біотопах, на снігу та льоду.
Систематичні ознаки відділу
Пігменти та запасні поживні речовини
До складу фотосинтетичних пігментів входять майже всі пігменти, властиві вищим рослинам: хлорофіли "а" та "b", всі відомі каротини (α, β, γ та ε- каротин), ксантофіли лютеїнового ряду (лютеїн, зеаксантин, неоксантин, віолаксантин, антераксантин). Специфічні ксантофіли, виявлені у сифонофіцієвих, ульвофіцієвих та празинофіцієвих водоростей - це сифонеїн, сифоноксантин, лороксантин, празиноксантин. У деяких празинофіцієвих виявлено також пігмент, подібний до хлорофілу "с".
Завдяки переважанню хлорофілів більшість видів Chlorophyta забарвлені у зелений колір. В екстремальних умовах (наприклад, у гіпергалінних водоймах) у деяких зелених водоростей "вмикається" механізм гіперсинтезу каротину та вторинних каротиноїдів, і тоді клітини набувають червоного забарвлення (Dunaliella salina). При старінні або при утворенні спочиваючих клітин у цитоплазмі може накопичуватись олія, забарвлена різними каротинами та ксантофілами (цю суміш називають гематохромом) у жовтий та червоний кольори.
Основним продуктом асиміляції є крохмаль. Він завжди відкладається в пластиді - або безпосередньо в стромі хлоропласта, або навколо піреноїда. Позапластидні асиміляти представлені олією (накопичується переважно при старінні клітин) та лейкозином. У сифонофіцієвих водоростей основним асимілятом може бути полісахарид інулін.
Цитологічні ознаки
Клітини зелених водоростей мають типово евкаріотичну будову і загалом подібні до клітин вищих рослин. Специфічні ознаки пов’язані, перш за все, з будовою клітинних покривів, хлоропласта, джгутикового апарату монадних клітин, особливостями мітозу та цитокінезу.
Клітинні покриви. Клітини переважної більшості видів зелених водоростей вкриті целюлозно-пектиновою оболонкою або оболонкою, каркас якої утворений кристалічно впорядкованими глікопротеінами, що збагачені гідроксипроліном. Значно рідше зустрічаються представники, позбавлені будь-яких покривів, окрім плазмалеми, тобто вони мають "голі" клітини. У зелених водоростей з класу празинофіцієвих, а також у монадних репродуктивних клітин деяких ульвофіцієвих та харофіцієвих водоростей на поверхні плазмалеми відкладаються від одного до кількох шарів субмікроскопічних органічних лусочок.
Ультратонка будова оболонки різноманітна і визначається систематичним положенням та віковими особливостями конкретних представників. Формування оболонки починається з розвитку т.зв. первинної оболонки, яка розвивається назовні від плазмалеми і складається з внутрішнього шару W1, кількох шарів т.зв. центрального триплету (шари W2 - W6), та у багатьох видів - з додаткового зовнішнього шару W7. Далі під первинною глікопотеіновою оболонкою може розвиватися вторинна оболонка, каркасну основу якої складають мікрофібрили целюлози, зцементовані пектиновими речовинами.
У багатьох одноклітинних зелених водоростей (зокрема, у Chlamydomonas reinhardtii та спорідених з ним родів) внутрішній шар W1 утворений нещільним шаром глікопротеінових мікрофібрил, що розташовуються більш-менш радіально. Далі розташовується центральний триплет, який зазвичай складається з трьох щільно прилеглих один до одного шарів - W2, W4, W6. Шар W2 являє собою мережу фібрил, орієнтованих паралельно до поверхні клітини (Woessner, Goodenough, 1993).
Проте все різноманіття структури оболонок є різними варіантами основного плану будови. Цей план найчіткіше простежується у молодих клітинах одноклітинних водоростей, оболонка яких складається з трьох шарів.
Перший, зовнішній шар дуже тонкий, утворений сульфатованими та кислими полісахаридами, і має торочкуватий або хвилястий край. Вважають, що він виконує рецепторні функції. Другий шар (т.з. первинна оболонка, періодично-глобулярний шар) є проміжним, утвореним упорядковано розташованими субодиницями-глобулами гідроксипролінів, глікопротеїнів та олігосахаридів. Третій шар (вторинна оболонка, фібрилярний шар) - внутрішній, він утворений мікрофібрилами целюлози, які "зцементовані" пектиновими речовинами. Вторинна оболонка найбільше розвинена у дорослих та старих клітинах. Під внутрішнім шаром знаходиться плазмалема.
У багатьох видів під час старіння клітин, утворення акінет та зигот зовнішній та проміжний шари можуть руйнуватися та зникати, а внутрішній, фібрилярний шар - потовщуватись та мультиплікуватись (рис. 24.1). Потовщення оболонки звичайно пов’язане з синтезом целюлозних мікрофібрил, збірка яких відбувається на поверхні плазмалеми особливими термінальними ферментативними комплексами. Тому оболонка завжди наростає зсередини назовні. Інколи у внутрішньому шарі є додаткові шари, утворені хітином або спорополеніном.
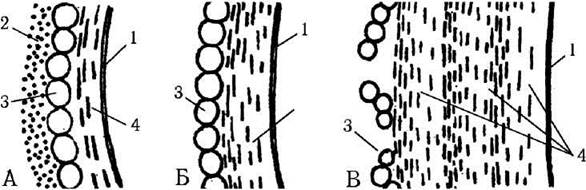
Рис. 24.1. Основний план будови клітинної оболонки зелених водоростей різного віку: А - молодих, Б - дорослих, В - старих клітин. 1 - плазмалема, 2 - зовнішній шар з сульфатованих та кислих полісахаридів, 3 - проміжний глобулярний глікопротеїновий шар, 4 - внутрішній фібрилярний целюлозний шар (схематизовано, за McLean, 1968, Константинова, 1991).
Клітинні оболонки бувають суцільними (у більшості видів) або складаються з кількох фрагментів (наприклад, з 2-х - у Microspora, 3-9 - у Treubaria); поверхня оболонок гладенька або несе різноманітні "прикраси" - шипи, шипики, ребра, бородавки та ін., інколи оболонки мають досить складний поровий апарат (особливо у кон’югат). У значної кількості видів оболонка мінералізується солями заліза, марганцю або кальцію.
Празинофіцієві водорості позбавлені оболонки; поверхня їх плазмалеми вкрита субмікроскопічними лусочками, що утворюються у цистернах комплексу Гольджі і виділяються назовні поблизу основи джгутиків.
Ядерний апарат зелених водоростей має типову евкаріотичну будову. Зовнішня мембрана ядерної оболонки ізольована, і не має структурного зв’язку з хлоропластами (рис. 24.2). У телофазі вона звичайно утворюється внаслідок злиття мембранних пухирців, що походять від комплексу Гольджі або ендоплазматичної сітки. Гомологічність цих структур досить часто можна спостерігати навіть в інтерфазі, коли зовнішня мембрана ядра переходить у канали ендоплазматичної сітки чи пов’язана з диктіосомами.
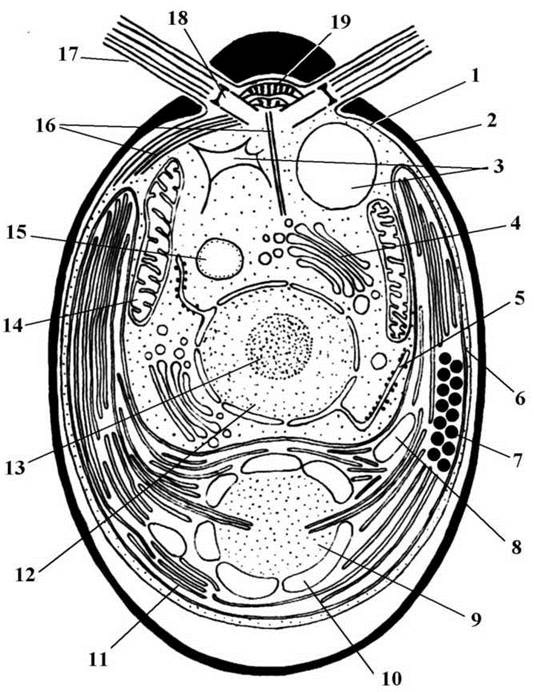
Рис. 24.2. Загальний план будови клітини зелених водоростей (на прикладі Chlamydomonas): 1 - плазмалема, 2 - клітинна оболонка, 3 - скоротливі ваколі, 4 - комплекс Гольджі, 5 - ендоплазматична сітка, 6 - двомембранна оболонка хлоропласту, 7 - стигма, 8 - строматичний крохмаль, 9 - піреноїд, 10 - крохмальна обгортка піреноїду, 11 - тилакоїди, зібрані у грану, 12 - ядро, 13 - ядерце, 14 - мітохондрія, 15 - краплина олії, 16 - мікротубулярні джгутикові корені, 17 - джгутик, 18 - базальне тіло джгутика, 19 - фібрилярне сполучне волокно базальних тіл (за Ettl, 1983).
В ядрі наявні одне-кілька ядерець, які (за винятком кон’югат) зникають під час мітозу. У монадних клітинах центриолі звичайно відсутні, їх функцію виконують базальні тіла джгутиків. У клітинах представників, позбавлених здатності до активного руху, центріолі наявні (за винятком кон’югат та харальних водоростей).
Мітоз закритий, напівзакритий або відкритий, може відбуватися у кількох варіантах, що розглядаються нижче, в описах окремих класів.
Ядерна ДНК за результатами аналізу різних генів найближче споріднена з ядерними ДНК глаукоцистофітових, родофітових, криптофітових та примнезіофітових водоростей, а також вищих рослин.
Фотосинтетичний апарат. Хлоропласти майже тотожні хлоропластам вищих рослин: вони одягнені двомебранною оболонкою, тилакоїди групуються у ламели по 3-6 або утворюють грани, оперезуючі тилакоїди відсутні. Звичайно у хлоропластах є один-кілька піреноїдів, переважно облямованих крохмалем.
У більшості монадних зелених водоростей у хлоропласті знаходиться стигма. Вона складається з кількох шарів осмієфільних, пігментованих каратиноїдами глобул, які розташовані безпосередньо під оболонкою хлоропласта. У місці розташування стигми оболонка хлоропласта тісно прилягає до плазмалеми. Вважають, що саме ця ділянка плазмалеми виконує функції фоторецептора, а стигма - функцію ширми, яка регулює кількість світла, що падає на фоторецептор. Із джгутиковим апаратом стигма зелених водоростей безпосередньо не пов’язана.
Хлоропластна ДНК замкнена у кільце, складається з 100-300 тис. пар нуклеотидів. Генофор розсіяний. За молекулярно-філогенетичними реконструкціями, хлоропластна ДНК зелених водоростей походить від ДНК синьозелених водоростей, її сестринськими лініями є хлоропластні ДНК глаукоцистофітових та червоних водоростей, а дочірньою - вищих рослин.
Вважають, що зелені водоростей є організмами з первинно- симбіотичнми хлоропластами. Останні утворилися внаслідок дивергенції фотосинтетичного апарату гіпотетичного фотоавтотрофного пращура, що набув пластиду внаслідок ендосимбіозу типу "гетеротрофний евкаріот + синьозелена водорість".
Мітохондріальний апарат представлений звичайно розгалуженим мітохондріоном з пластинчастими кристами. Мітохондріальна ДНК складається з 16-80 тис. пар нуклеотидних основ (у морських сифональних водоростей - до 220 тис. пар), і за молекулярно-філогенетичними реконструкціями близька до мітохондріальної ДНК інших евкаріот, що мають мітохондрії з пластинчастими кристами.
Джгутикові стадії. Переважна більшість монадних клітин зелених водоростей має по два ізоконтні та ізоморфні джгутики. Рідше зустрічаються чотири- або багатоджгутикові (стефаноконтні) форми (рис. 24.3). Серед примітивних Chlorophyta є види з одним, двома-трьома неоднаковими, шістьма або вісьмома однаковими за довжиною джгутиками.
Поверхня джгутиків звичайно гладенька або вкрита органічними субмікроскопічними лусочками, які формуються в особливих резервуарах комплексу Гольджі. У деяких родів наявні прості трубчасті мастигонеми.
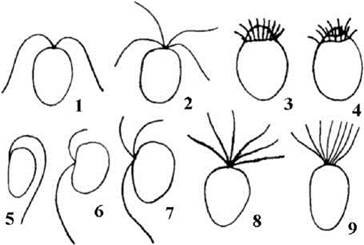
Рис. 24.3. Поширені (1-4) та відносно рідкісні (5-9) плани будови монадних стадій зелених водоростей: 1, 2- дво- та чотириджгутиковий ізоконтний, 3, 4 - стефаноконтний з кільчастим (3) та спіральним (4) розміщенням джгутиків, 5 - одноджгутиковий, 6, 7 - дво- та триджгутиковий гетероконтний, 8, 9 - шести- та восьмиджгутиковий ізоконтний.
Перехідна зона джгутика має специфічну зірчасту структуру, що виявлена тільки в монадних клітинах зелених водоростей та вищих рослин (рис. 24.4). Крім зірчастої структури, у примітивних зелених водоростей з класу празинофіцієвих у перехідній зоні можуть бути спіралеподібна структура та проміжна платівка.
Система джгутикових коренів складається з мікротубулярних та мікрофібрилярних коренів. Плани організації кореневої системи у різних класах відділу різна.
Інші органели. Вакуолярний апарат представлений справжніми вакуолями з клітинним соком - тонопластом. У клітинах багатьох монадних та гемімонадних прісноводних та наземних водоростей звичайно є дві, зрідка - кілька скоротливих вакуолей. Деяким празинофіцієвим водоростям властиві трихоцисти та слизові тільця, що за будовою нагадують відповідні структури криптофітових та рафідофітових водоростей.
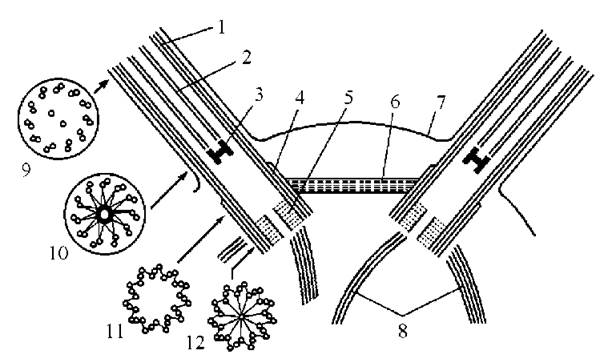
Рис. 24.4. Загальний план будови джгутикового апарату зелених водоростей: 1 - периферичні мікротрубочки аксонеми, 2 - центральні мікротрубочки, 3 - зірчаста структура у перехідній зоні, 4 - базальне тіло, 5 - нижня частина базального тіла - "візкове колесо", 6 - верхнє сполучне волокно базальних тіл, 7 - плазмалема, 8 - мікротубулярні джгутикові корені, 9-12 - структура аксонеми (9), перехідної зони з зірчастою структурою (10), базального тіла у верхній частині (11) та зоні "візкового колеса" (12).
Типи структури тіла
За морфологією Chlorophyta є найрізноманітнішим відділом водоростей. Зелені водорості бувають як мікро- так і макроскопічними, одно- та багатоклітинними, деякі мають неклітинну (сифональну або сифонокладальну) будову. В межах відділу представлені майже всі типи морфологічної структури тіла - монадний, гемімонадний, кокоїдний, сарциноїдний, трихальний, гетеротрихальний, паренхіматозний, сифональний та сифонокладальний. Тип структури є однією з головних ознак при поділі зелених водоростей на порядки за класичними системами. Як одно-, так і багатоклітинні водорості можуть утворювати різноманітні колонії, вести вільний або прикріплений спосіб життя.
Розмноження та ріст багатоклітинних таломів
У зелених водоростей спостерігаються усі основні способи нестатевого розмноження і статевого процесу, відомі у евкаріотичних водоростей. В цілому, у межах відділу спостерігається тенденція заміни рухливих репродуктивних клітин нерухомими, та редукція нестатевого розмноження спорами за рахунок інтенсифікації вегетативного та статевого розмноження. В окремих гілках, що становлять тупий кут еволюції (наприклад, хлорококальні або сценедесмальні) наявна тенденція вторинної втрати статевого процесу та перехід до апогамії.
У зелених водоростей відомі клітинні поділи за типами схізотомії, схізогонії та цитотомії (рис. 24.5). Два останніх типи спричинюють ріст багатоклітинних таломів, причому цитотомія за участю клітинної платівки зустрічається тільки у відділі Chlorophyta.
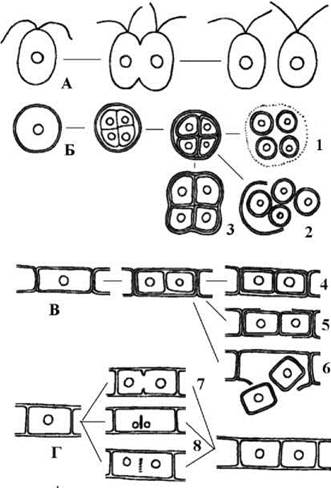
Рис. 24.5. Способи клітинного поділу у зелених водоростей: А - схізотомія, Б, В - схізогонія, Г - цитотомія (1, 2, 6 - схізогонія, наслідком якої є споруляція; 3 - схізогонія, що призводить до
утворення та росту багатоклітинних сарциноподібних агрегатів; 4, 5 - схізогонія, що спричинює ріст таломів у нитчастих та гетеротрихальних форм; 7, 8 - цитотомія за участю кільцевої борозни та клітинної платівки, яка завжди обумовлює наростання талому).
Життєві цикли
Зеленим водоростям властиві різні типи життєвих циклів. Близько чверті відомих видів є агамними або апогамними, з життєвими циклами за типом цикломорфозу.
До еугамних належить більшість видів відділу, і їх життєві цикли представлені гаплофазними, гаплодиплофазними та диплофазними циклами з зиготичною, гаметичною, споричною та соматичною редукцією, з ізо- та гетероморфною зміною поколінь. Життєві цикли можуть бути однотипними у всіх представників таксону рангу класу або порядку, а можуть відрізнятись навіть у різних видів одного роду.
Система зелених водоростей
Зелені водорості - це об’єкт інтенсивних досліджень. Великий обсяг нової інформації, необхідність її узагальнення та систематизації призводять до того, що таксономія Chlorophyta перебуває у надзвичайно динамічному стані.
Сьогодні існує багато систем зелених водоростей. Згідно з засадами, на яких вони ґрунтуються, всі ці системи можна поділити на: а) класичні морфолого-онтогенетичні, б) цитологічні, в) молекулярно-філогенетичні системи.
Морфолого-онтогенетичні системи
Системи даного типу побудовані на засадах, розроблених на початку ХХ століття видатним альгологом Адольфом Пашером. Поділ зелених водоростей на таксони високого рангу (класи, порядки) проводять, перш за все, у відповідності з типами морфологічних структур та особливостями статевого процесу. Найбільшого визнання серед подібних систем дістала система Б. Фотта (Fott, 1971, 1973) та пізніші її модифікації. Зокрема, в Україні широкого використання набула система, викладена у навчальному посібнику "Пресноводные водоросли Украинской ССР" (Топачевский, Масюк, 1984).
Класичні системи є штучними, вони не відображають справжніх родинних зв’язків таксонів, і тому непридатні для вирішення ряду завдань (наприклад, у галузі порівняльної флористики, історичного аналізу, філогенії). Проте вони мають кілька переваг над молодшими цитологічними та молекулярно-філогенетичними системами: класичні системи зручні для ідентифікації конкретних родів та видів, для визначення місця у системі нових таксонів. Дуже важливим є й те, що сучасні визначники зелених водоростей базуються поки що саме на класичних системах.
Згідно з класичними морфолого-онтогенетичними системами, відділ Chlorophyta поділяється на три класи - Chlorophyceae (або Euchlorophyceae), Conjugatophyceae та Charophyceae. В основу поділу на класи покладено тип статевого процесу, наявність або відсутність джгутикових стадій в онтогенезі, будову статевих органів. Поділ класів на порядки проводять, в першу чергу, за типами морфологічної структури тіла.
Клас Chlorophyceae об’єднує одно-, багато- та неклітинні водорості, здатні до нестатевого розмноження за допомогою спор та/або до хологамного чи мерогамного статевого розмноження. Клас поділяють на 11 порядків. Нижче подано їх основні ознаки та приклади характерних представників.
Порядок Pedinomonadales - монадні водорості з нетиповою будовою джгутикового апарату (одноджгутикові, дво- або триджгутикові гетероконтні види). Характерні представники - Pedinomonasта Monomastix.
Порядок Volvocales - монадні водорості з типовою будовою джгутикового апарату. Характерні представники - Dunaliella, Chlamydomonas, Haematococcus, Gonium, Pandorina, Volvox.
Порядок Tetrasporales - гемімонадні водорості, наприклад, Tetraspora.
Порядок Chlorococcales - кокоїдні водорості. Характерні представники - Chlorococcum, Trebouxia, Hydrodictyon, Chlorella, Scenedesmus, Ankistrodesmus.
Порядок Chlorosarcinales - сарциноїдні водорості. Характерні представники - Chlorosarcinopsis, Tetracystis, Chlorokybus.
Порядок Ulotrichales - трихальні водорості з дво- або чотириджгутиковими репродуктивними клітинами. Характерні представники - Ulothrix, Uronema, Klebsormidium, Stichococcus, Raphidonema, Ulva, Enteromorpha, Microspora.
Порядок Chaetophorales - гетеротрихальні водорості з дво- або чотириджгутиковими репродуктивними клітинами. Характерні представники - Stigeoclonium, Chaetophora, Draparnaldia, Pleurastrum, Coleochaete.
Порядок Oedogoniales - трихальні та гетеротрихальні водорості з стефаноконтними репродуктивними клітинами та особливим типом клітинного поділу з утворенням ковпачків. Характерні представники - Oedogonium, Bulbochaete.
Порядок Bryopsidales - сифональні водорості. Характерні представники - Bryopsis, Codium, Acetabularia.
Порядок Siphonocladales - сифонокладальні водорості, що мають ізо- або гетерогамний статевий процес. Характерні представники - Rhizoclonium, Cladophora, Siphonocladus.
Порядок Sphaeropleales - сифонокладальні водорості, що мають оогамний статевий процес. Зигота при розвитку проростає в особливий багатоядерний заросток. Характерний представник - Sphaeroplea.
Клас Conjugatophyceae об’єднує кокоїдні та трихальні водорості зі статевим процесом за типом кон’югації. Будь-які монадні стадії в циклі розвитку відсутні. За типами структури клас поділяють на 4 порядки.
Порядок Mesotaeniales - кокоїдні водорості з гладкою оболонкою, у яких клітина не диференційована на напівклітини. Характерні представники - Mesotaenium, Cylindrocystis, Spirotaenia.
Порядок Gonatozygales - трихальні водорості з нетривкими нитками та скульптурованими оболонками. Характерний представник - Gonatozygon.
Порядок Desmidiales - кокоїдні водорості зі скульптурованою оболонкою та складним поровим апаратом. Клітини переважно диференційовані на напівклітини. Характерні представники - Penium, Closterium, Cosmarium, Micrasterias.
Порядок Zygnematales - трихальні водорості з гладкими оболонками. Характерні представники - Spirogyra, Zygnema, Mougeotia.
Клас Charophyceae об’єднує гетеротрихальні водорості специфічної членисто-мутовчастої будови, у яких статевий процес - високоспеціалізована оогамія, а джгутикові стадії представлені лише сперматозоїдами. Включає один порядок - Charales. Характерні представники - Chara та Nitella.
Цитологічні системи
У 70-і роки електронно-мікроскопічні дослідження показали, що у зелених водоростей наявні два принципово відмінні типи організації джгутикових коренів - хрестоподібна та асиметрична однобічна, три типи цитокінезу - за участю кільцевої борозни, фікопласту або фрагмопласту, три типи мітозу - закритий, напівзакритий та відкритий, кілька різних варіантів будови веретена поділу, різні типи поведінки хромосом у метафазі, різні варіанти ультратонкої будови клітинних покривів та ін. Виходячи з наявності такої різноманітності ультраструктурних ознак, одразу кілька альгологів, що працювали у галузі порівняльної цитології зелених водоростей - К.Д. Стьюарт, К.Р. Меттокс, М. Шадефо, Т. Христенсен, та ін., - спробували перебудувати систему зелених водоростей на цитологічній основі. Відділ зелених водоростей був розділений на класи Pedinophyceae, Micromonadophyceae, Prasinophyceae, Chlorophyceae, Ulvophyceae та Charophyceae.
Пізніше система класів постійно деталізувалась, вводилися нові ознаки класів, самі класи подрібнювались і їх кількість зростала1. Наприклад, у найсучаснішому підручнику К. ван ден Хука, Д. Манна та Х. Джанса (van den Hoek, Mann, Jahns, 1995), за ультраструктурними ознаками Chlorophyta розділено на 11 класів. Проте таксономічна вага тих чи інших ультраструктурних ознак лишалась проблематичною. Ситуація почала суттєво змінюватись у 90-х роках ХХ сторіччя.
Молекулярно-біологічні системи
Для розв'язання питання стосовно того, які саме ознаки найреальніше відображають основні напрямки еволюції зелених водоростей (і, відповідно, які варіанти системи Chlorophyta найбільш наближені до природньої системи), у другій половині 80-х років почали застосовувати молекулярно-біологічні методи. Дослідження спочатку ґрунтувалися переважно на аналізі нуклеотидних послідовностей ядерного гену, що кодує 5S та 18S субодиниці рибосомальної РНК. Пізніше аналогічні роботи стали виконувати й на інших генах, причому не тільки ядерних, але й хлоропластних та мітохондріальних (наприклад, генах фактору подовження тубуліну, цитохрому "с", актину, RUBISCO і т.д.). Для аналізу результатів сіквенсів нуклеотидних послідовностей певних генів на початку 90-х років були розроблені численні варіанти побудови молекулярно-філогенетичних дерев та оцінки їх інформативності.
Вже перші філогенетичні дерева, побудовані на базі молекулярних даних, показали монофілетичність Chlorophyta, гетерогенність традиційного класу Chlorophyceae, ширший обсяг класу Charophyceae (порівняно з традиційним) та його філогенетичний зв'язок з вищими рослинами, а також необхідність виділення кількох додаткових класів. Отримані результати дозволили також провести процедуру зважування різних морфологічних та цитологічних ознак та виділити серед них ті, що узгоджувались з молекулярно- філогенетичними реконструкціями. Наприкінці 90-х років було підведено підсумки дискусії про основні напрямки еволюції Chlorophyta та розроблено систему класів зелених водоростей, найбільш наближену до природньої системи.
Молекулярно-філогенетичні реконструкції, з одного боку, підтвердили високу філогенетичну вагу таких цитологічних ознак, як будова джгутикового апарату, особливості мітозу та цитокінезу, а також показали, що в межах Chlorophyta сьогодні існує 7 великих і чітко окреслених груп таксонів, які слід розглядати як самостійні класи. Цікавим виявився і той факт, що ці класи узгоджувались не тільки з цитологічними ознаками, але й з екологічними особливостями видів, які до них увійшли.
Згідно до молекулярно-філогенетичних реконструкцій, зелені водорості поділяють на класи Prasinophyceae, Chlorophyceae, Trebouxiophyceae, Ulvophyceae, Siphonophyceae, Zygnematophyceae(=Conjugatophyceae) та Charophyceae. Огляд ознак, що лежать в основі цього поділу, подано нижче.
Систематичні ознаки класів зелених водоростей
Основні типи клітинних покривів
Плазмалема у зелених водоростей може бути вкрита двома принципово відмінними типами клітинних покривів - або субмікроскопічними лусочками, що утворюються у комплексі Гольджі, або целюлозно-пектиновою оболонкою, фібрили якої синтезується на поверхні плазмалеми, наприклад, за участю термінальних ферментних комплексів. Перший тип покривів властивий вегетативним клітинами празинофіцієвих водоростей, другий - представникам всіх інших класів. Цікаво, що у харофіцієвих та ульвофіцієвих водоростей зооспори та гамети також вкриті субмікроскопічними лусочками, подібними до лусочок празинофіцієвих водоростей, хоча вегетативні клітини одягнені клітинною оболонкою. Згідно з біогенетичним законом, це може свідчити про походження Charophyceae та Ulvophyceae від гіпотетичних джгутикових предків, подібних до сучасних Prasinophyceae.
Типи кореневих систем джгутиків
У спрощеному вигляді, кореневі системи джгутиків зелених водоростей представлені або хрестоподібним, або асиметричним типом.
У водоростей з хрестоподібною кореневою системою від базальних тіл джгутиків відходять чотири хрестоподібно розташовані мікротубулярні корені. Супротивні корені мають однакову кількість мікротрубочок. Базальні тіла джгутиків з'єднуються між собою сполучним волокном. У деяких представників вглиб клітини також спрямовані два ризопласти.
В узагальненому вигляді варіанти хрестоподібної системи джгутикових коренів позначають формулою вигляду n1-n2-n1-n2, де n1 - кількість
мікротрубочок першого та третього коренів, а n2 - кількість мікротрубочок другого та четвертого коренів. Для переважної більшості видів з хрестоподібною системою ця формула має вигляд (5-6)-2-(5-6)-2, тобто кількість мікртрубочок першого та третього коренів дещо варіює, а другий та четвертий корені зазвичай складаються з двох мікротрубочок. Відомі також інші варіанти. Наприклад, у трентеполіальних водоростей формула має вигляд 6-4-64, у ульвовальних та кладофоральних - 3-2-3-2.
За орієнтацією базальних тіл хрестоподібні кореневі системи поділяють на три типи - з базальними тілами зміщеними за годинниковою стрілкою (1-7 система), проти годинникової стрілки (11-5 система) або супротивними (12-6 система). Для визначення типу орієнтації передню частину клітини розглядають у поперечній площині, джгутики орієнтують вертикально. Якщо після цього через центри базальних тіл провести вісь, то години циферблату, через які вона проходить, будуть відповідати орієнтації базальних тіл (рис. 24.6).
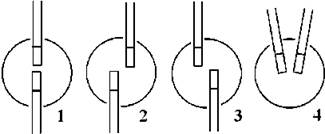
Рис. 24.6. Орієнтація базальних тіл у зелених водоростей: 1 - супротивна (12-6), 2 - зміщена за годинниковою стрілкою (1 -7), 3 - зміщена проти годинникової стрілки (11-5), 4 - однобічна (при асметричній кореневій системі).
Системи типу 12-6 та 1-7 характерні для водоростей з класу Chlorophyceae, система 11-5 - для Trebouxiophyceae, Ulvophyceae та Siphonophyceae.
У водоростей з асиметричною кореневою системою від базальних тіл джгутиків вглиб клітини відходить лише один мікротубулярний корень, побудований з багатьох (до кількох десятків) мікротрубочок. З базальними тілами він пов"язаний через багатошарову мікрофібрилярну структуру. Монадні клітини з таким типом кореневої системи звичайно дорзовентральні, джгутики спрямовані у вентральний бік, а базальні тіла розташовуються більш-менш паралельно. Така система характерна для монадних клітин представників Charophyceae та вищих рослин.
У водоростей з класу Prasinophyceae кореневі системи можуть бути як хрестоподібними, так і асиметричними, або утворювати комбінований варіант - хрестоподібну систему з додатковим асиметричним коренем.
Крім типів симетрії (хрестоподібна чи асиметрична), різноманітність кореневих систем зелених водоростей простежується також за будовою мікрофібрилярних коренів, сполучних волокон базальних тіл, наявністю термінальних ковпачків на базальних тілах, за кутами нахилу базальних тіл одне до одного та ін.
Типи мітозу
Різноманітність мітозу у зелених водоростей простежується, в першу чергу, за його перебігом на стадіях профази та телофази.
У профазі ядерна оболонка або лишається інтактною або перфорується чи зникає. Всі ці типи поведінки ядерної оболонки відомі у Prasinophyceae; у Chlorophyceae, Ulvophyceae та Siphonophyceae мітоз закритий, у Conjugatophyceae та Trebouxiohyceae - напівзакритий, а у Charophyceae - відкритий.
У відповідності з поведінкою у телофазі, веретено поділу розглядають або як те, що зберігається протягом всієї телофази - т.з. персистентне, або як зникаюче - колапсуюче. У випадку персистентного веретена дочірні ядра відходять одне від одного на значну відстань, тоді як при колапсуючому веретені лишаються поруч. Персистентне веретено властиве представникам Ulvophyceae, Siphonophyceae, Zygnematophyceae, Charophyceae, а зникаюче - Chlorophyceae та Trebouxiophyceae. Представники празинфіцієвих водоростей можуть мати веретено як зникаюче, так і персистентне.
Типи цитокінезу
Поділ клітини, який відбувається по закінченні поділу ядра, здійснюється кільцевою борозною або шляхом утворення клітинної платівки.
Кільцева борозна. При цьому типі поділу плазмалема і внутрішній шар оболонки вростають у середину клітини, перекриваючи її подібно до ірисової діафрагми. Вростання оболонки пов’язане з доцентровим відкладанням целюлозних мікрофібрил, що траспортуються у сплощених пухирцях комплексу Гольджі. Поділ за участю кільцевої борозни зустрічаєься в усіх класах зелених водоростей, за винятком Siphonophyceae, у яких цитокінез відсутній, оскільки представники цього класу мають сифональну організацію.
Клітинна платівка, навпаки, формується відцентрово. В зоні майбутньої клітинної перегородки утворюється система паралельних мікротрубочок, що є організатором клітинної платівки. Мікротрубочки організатора клітинної платівки виконують функції направляючих рейок, по яким до місця утворення перегородки рухаються пухирці комплеку Гольджі, що містять мікрофібрили целюлози. Залежно від орієнтації мікротрубчок по відношенню до майбутньої клітинної перегородки, систему організатора клітинної платівки поділяють на два типи - фікопласт або фрагмопласт.
Якщо мікротрубочки організатора орієнтуються паралельно до майбутньої перегородки, то кажуть, що вони утворюють фікопласт. У монадних форм мікротрубочки фікопласта утворюються за участю першого та третього джгутикових коренів. У нерухомих клітин фікопласт утворюється на початку телофази з центральної зони веретена внаслідок часткового руйнування та переорієнтації його мікротрубочок. Вважають, що зникаючий тип веретена є результатом використання його мікротрубочок на побудову фікопласта. Фікопласт є характерним для хлорфіцієвих та требуксіофіцієвих водоростей. Він виявлений також у деяких представників Prasinophyceae. Цікаво, що у останніх внаслідок злиття всередині фікопласту мембранних пухирців утворюється не клітинна платівка, а лише частини плазмалеми дочірніх клітин, оскільки пухирці не містять целюлозного матеріалу.
Коли мікротрубочки організатора клітинної платівки орієнтуються перпендикулярно до майбутньої перегородки, то таку систему мікротрубочок називають фрагмопластом. Мікротрубочки фрагмопласту синтезуються у телофазі поміж мікротрубочками центральної частини персистентного веретена. Фрагмопласт містить також мікрофібрили актину. Мікротрубочки та мікрофібрили фрагмопласта направляють та вирівнюють у центральній зоні веретена пухирці комплексу Гольджі з матеріалом майбутньої клітинної перегородки. Фрагмопласт виявлений у водоростей з Conjugatophyceae та Charophyceae, а також у трентеполіальних водоростей та вищих рослин (табл. 24.1).
Таблиця 24.1. Деякі цитологічні ознаки класів зелених водоростей
Класи |
Клітинні покриви |
Коренева система джгутиків |
Мітоз |
Веретено |
Організатор клітинної платівки |
Prasino- |
лусочки |
хр., асим., хр.+асим. |
відкритий, закритий |
персистентне, колапсуюче |
(фікопласт) |
Chloro- |
кл.обол. |
хр. (1-7 або 12-6) |
закритий |
колапсуюче |
фікопласт |
Trebouxio |
кл.обол. |
хр.(11-5) |
напівзакр. |
колапсуюче |
фікопласт |
Ulvo- |
кл.обол. |
хр.(11-5) |
закритий |
персистентне |
- |
Siphono- |
кл.обол. |
хр.(11-5) |
закритий |
персистентне |
- |
Charo- |
кл.обол. |
асим. |
напівзакр., відкритий |
персистентне |
фрагмопласт |
Zygnemato- |
кл.обол. |
- |
напівзакр. |
персистентне |
фрагмопласт |
Скорочення: кл.обол. - клітинна оболонка, хр. - хрестоподібна, асим. - асиметрична, напівзакр. - напівзакритий.
Додаткові ознаки
З основними ознаками класів певним чином корелює кілька додаткових - особливості пігментного складу та продуктів асиміляції, здатність до утворення трихоцист та слизових тіл, типи морфологічної структури тіла, екологічні особливості, та ін.
Найважливіші цитологічні ознаки класів зелених водоростей подані у таблиці 24.1.
24.1. Вихідні лінії: клас Prasinophyceae
Об’єднує біля 200 видів одноклітинних, здебільшого монадних водоростей, що мешкають переважно у морях. Празинофіцієві вважаються найдавнішими зеленими водоростями, що дали початок усім лініям Chlorophyta. На відміну від інших класів, для Prasinophyceae характерні різноманітні плани будови клітин, різні типи кореневих систем джгутиків, мітозу та цитокінезу. Викопні рештки, що нагадують цисти сучасних празинофіцієвих водоростей, датуються віком біля 1.2 млрд років.
Ознаки та характеристика класу
Біохімічні ознаки
Хлоропласти всіх празинофіцієвих водоростей містять хлорофіли а та b. Крім того, у десяти видів виявлено пігмент, що нагадує хлорофіл с. Цей пігмент, на думку одних дослідніків, є справжнім хлорофілом с, на думку інших, подібним, проте не ідентичним до хлорофілу с пігментом - складним монометиловим ефіром Mg-вмісного 2,4-дивінілфеопорфірину а5. У деяких видів замість лютеїну знайдено празиноксантин. Відомі також представники, у яких наявні ксантофіли, характерні для ульвофіцієвих та сифонофіцієвих водоростей - сифонеїн та сифоноксантин.
Цитологічні особливості
Клітинні покриви. Празинофіцієві водорості з порядку Pedinomonadales мають клітини, вкриті лише плазмалемою. Проте у більшості празинофіцієвих над плазмалемою розташовується один-кілька шарів субмікроскопічних лусочок з органічних речовин. Інколи лусочки щільно з'єднуються одна з одною, формуючи т.з. теку, яка у світловому мікроскопі виглядає як клітинна оболонка (рис. 24.7).
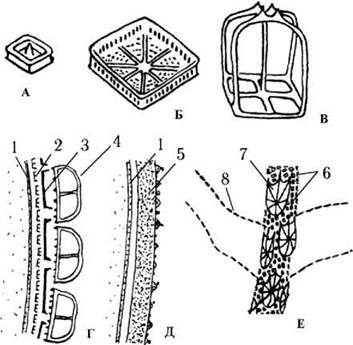
Рис. 24.7. Типи лусочок та будова клітинних покривів празинофіцієвих водоростей: А-В - клітинні лусочки (А - підстилаючі, Б - проміжні, В - поверхневі); Г - розташування шарів лусочок на поверхні клітин Pyramimonas; Д - тека у Tetraselmis; Е - джгутикові лусочки у Pyramimonas. 1 - плазмалема, 2-4 - клітинні лусочки: підстилаючі (2), проміжні (3), поверхневі (4); 5 - тека, що утворилася внаслідок злиття підстилаючих лусочок, 6-8 - джгутикові лусочки: підстилаючі (6), проміжні (7), лусочкові волоски (8) (схематизовано за Sym, Pienaar, 1991, 1993; Lewin, Lee, 1985).
Лусочки залежно від їх розмірів та морфології поділяють на три основні типи: підстилаючі, проміжні та поверхневі. Підстилаючі лусочки дуже дрібні (до 50 нм завдовжки), сплощені, мають переважно прямокутну форму. Проміжні лусочки більші (250-350 нм завд.), різних обрисів, проте звичайно з високим краєм, через що нагадують відкриті коробочки. Поверхневі лусочки ще більші (до 600-700 нм завд.), як правило, високі, інколи з шипиками та щетинками, мають складну архітектуру.
Лусочки утворені вуглеводами із незначною (до 5%) домішкою білків. Основними вуглеводами є пектиноподібні карбогідрати на основі 2-кето- цукрових кислот з домішкою нейтральних цукрів типу арабінози, галактози, глюкози, ксилози. Наявність у складі клітинних покривів 2-кето-цукрових кислот вважається специфічною особливістю празинофіцієвих водоростей; крім Prasinophyceae, ці кислоти виявлено також у вищих рослин та деяких бактерій.
Тека за хімічним складом дещо відрізняється від лусочок: вона містить не тільки 2-кето-цукрові кислоти, але й значну кількість галактуронової кислоти, іонів кальцію та сульфатних залишків. Саме галактуронова кислота зв’язує дрібні субмікроскопічні лусочки у єдиний цілісний клітинний покрив.
Утворення лусочок відбувається у пухирцях комплексу Гольджі, які надалі мігрують до спеціального резервуару. Лусочковий резервуар відкривається отвором при основі джгутиків, через цей отвір лусочки виходять назовні і відкладаються на плазмалемі. У представників, що мають теку, лусочки не накопичуються у резервуарі, а безпосередньо у пухирцях комплексу Гольджі об'єднуються у фрагменти теки.
Деякі празинофіцієві за певних умов виділяють назовні шар слизу і переходять у пальмелевидний стан. У переважній більшості випадків такий слиз є аморфним, проте у видів роду Prasinocladusвін утворює слизові трубки.
За несприятливих умов та залежно від фаз місячного циклу частина празинофіцієвих водоростей формує цисти, вкриті особливою двошаровою оболонкою. Зовнішній шар утворений спорополеніном із досить значною кількістю ліпоїдів, він може бути гладеньким або скульптурованим субмікроскопічними лусочками, інколи - утворювати ребра або вирости. Внутрішній шар пектиновий, із залишками сульфатованої галактуронової кислоти. Під час проростання цист внутрішній шар звичайно повністю ослизнюється, через що дочірні клітини виходять назовні у слизовому міхурі.
Ядерний апарат та особливості мітозу. У празинофіцієвих водоростей справжні центриолі відсутні, а їх функції опосередковано виконують базальні тіла джгутиків. Між базальними тілами та ядром у більшості видів існує структурний зв'язок, що реалізується за допомогою ризопластів (рис. 24.8).
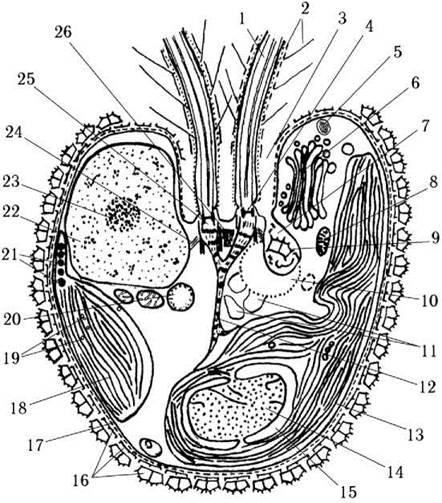
Рис. 24.8. Схема будови клітини празинофіцієвої водорості (на прикладі Pyramimonas): 1 - джгутик, 2 - джгутикові лусочки, 3 - апікальна заглибина, 4 - перехідна зона джгутику із зірчастою структурою, 5 - трихоциста, 6 - комплекс Гольджі, 7 - лусочковий резервуар, 8 - мітохондріальний профіль, 9 - лусочка у резервуарі, 10 - скоротлива вакуоля, 11 - мікротільця, 12 - ризопласт, 13 - хлоропласт, 14 - піреноїд, 15 - крохмальна обгортка піреноїда, 16 - клітинні лусочки, 17 - плазмалема, 18 - тилакоїд, 19 - гранули строматичного крохмалю, 20 - краплина олії, 21 - стигма, 22 - ядро, 23 - ядерце, 24 - мікротубулярний корінь, 25 - базальне тіло, 26 - синістосома (схематизовано за Ettl, 1983; Inouye, Hori, Chihara, 1983).
На початку мітозу джгутики або лишаються інтактними, або скидаються, проте ніколи не втягуються. Через це на початкових стадіях каріокінезу у клітині практично немає запасу вільного тубуліну. Базальні тіла реплікуються, проте не мігрують углиб клітини. Ризопласти частково руйнуються у зоні, що безпосередньо прилягає до ядерної оболонки. Тут спостерігається концентрація гранулярного матеріалу, з якого починає формуватися веретено поділу. Цьому процесу передує часткова дезінтеграція мікротубулярних коренів, внаслідок чого у цитоплазмі з’являється необхідна для побудови веретена кількість тубуліну.
Ядерна оболонка під час мітозу може лишатися інтактною (наприклад, у Tetraselmis), або зникати (Pyramimonas). Веретено поділу або персистентне (Nephroselmis), або швидко руйнується у телофазі (Tetraselmis).
Після закінчення каріокінезу між дочірніми ядрами скупчуються пухирці комплексу Гольджі, внаслідок злиття яких у доцентровому напрямку росте поздовжня кільцева борозна. У деяких видів цей процес відбувається за участю фікопласту (рис. 24.9).
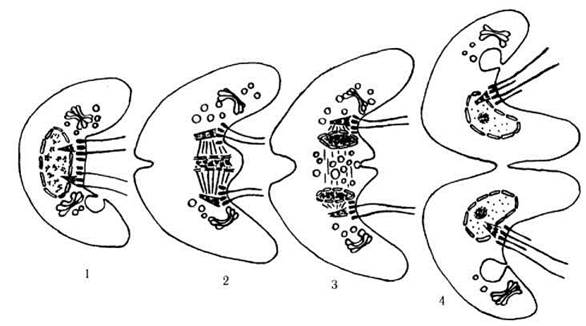
Рис. 24.9. Мітоз у Pyramimonas: 1 - рання профаза, 2 - метафаза, 3 - пізня телофаза, 4 - рання інтерфаза (схематизовано за Woods, Triemer, 1981).
Таким чином, празинофіцієвим водоростям властиві закриті та відкриті мітози з персистентним або зникаючим веретеном, ризопластами, що виконують функції центру організації мікротрубочок веретена, та цитокінезом, що відбувається за допомогою кільцевої борозни за участю комплексу Гольджі з формуванням фікопласту або без нього.
Джгутиковий апарат Prasinophyceae вельми різноманітний. У переважної більшості видів джгутики виходять з апікальної або латеральної заглибини - т.з. "джгутикової ямки", що може розглядатися як редукована глотка. Деякі представники мають джгутики, що відходять від опуклої поверхні клітини. Кількість джгутиків варіює від одного до восьми, вони можуть бути як ізоконтними, так і гетероконтними.
Гетероконтність спостерігається у тих видів, які на початку поділу не скидають джгутики, а рівномірно розподіляють їх між дочірніми клітинами. У цьому випадку кожна дочірня клітина отримує половину джгутиків від материнської клітини, а решту будує de novo. Процес збірки нового джгутика у празинофіцієвих водоростей, на відміну від водоростей інших класів, відбувається повільно, оскільки запасу вільного тубуліну у цитоплазмі немає. Тому джгутики, отримані у спадок від материнської клітини довші, ніж джгутики, що синтезуються заново. Перші в інтерфазі звичайно вже не ростуть, тоді як довжина останніх постійно збільшується пропорційно до кількості синтезованого клітиною тубуліну.
Якщо перед поділом материнська клітина скидає джгутики, тобто відбувається автотомія, то дочірні клітини синтезують de novo всі джгутики. В цьому випадку монадні клітини ізоконтні.
Джгутики вкриті звичайно одним-трьома шарами субмікроскопічних лусочок, що утворюються в комплексі Гольджі і накопичуються в тому ж резервуарі, що й клітинні лусочки. Внутрішній шар утворений дрібними (40-70 нм завширшки) сплощеними прямокутними лусочками, далі розташований проміжний шар більших (200-400 нм) лусок з шипами; зовнішній шар складають лусочки зі щетинками до 1 мкм завдовжки. Кількість шарів, морфологію та хімічний склад джгутикових лусочок використовують як систематичні ознаки на рівні порядків.
У перехідній зоні джгутика празинофіцієві водорості мають характерну для зелених водоростей зірчасту структуру. У багатьох видів є також додаткові елементи перехідної зони - одна чи дві платівки, спіралеподібна структура або циліндр (рис. 24.10).
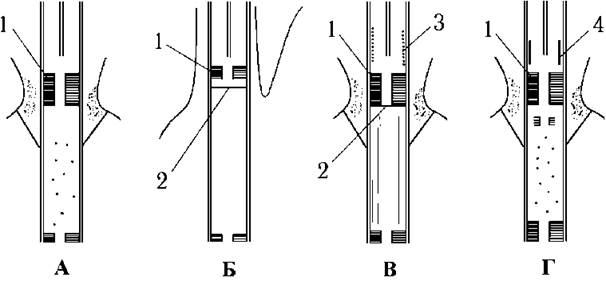
Рис. 24.10. Перехідна зона джгутика празинофіцієвих водоростей у поздовжньому розрізі: А - Mesostigma, Б - Mantoniella, В - Pterosperma, Г - Pyramimonas (1 - зірчаста структура, 2 - поперечна платівка, 3 - спіралеподібна структура, 4 - циліндр) (згідно до Sym, Pienaar, 1993).
Базальні тіла джгутиків, на відміну від базальних тіл у представників інших класів, вельми видовжені. Звичайно вони з'єднуються з плазмалемою та інколи з ядром за допомогою двох поперечно-смугастих ризопластів, побудованих з білка центрину. Ризопласти здатні скорочуватись, через що клітини можуть змінювати форму. Скорочення відбувається внаслідок спіралізації фібрил центрину. Цей процес здійснюється спонтанно і тільки за наявності іонів кальцію. Протилежний процес - деспіралізація центрину - навпаки, потребує витрат енергії і відбувається за участю АТФ.
Між собою базальні тіла пов’язані різноманітними мікрофібрилярними сполучними структурами. Так, у мамієлальних водоростей базальні тіла з’єднані одним гладеньким центриновим волокном. У псевдоскурфієльдіальних водоростей таких волокон два - поперечно-смугасте знаходиться одразу під перехідною зоною джгутика (т.з. дистальне волокно), гладеньке - трохи нижче (проксимальне волокно). У пірамімонадальних водоростей і дистальні, і проксимальні волокна гладенькі, а крім них між базальними тілами розташовується масивна поперечно-смугаста структура, що отримала назву синістосоми. У водоростей, що мають теку, крім дистальних та проксимальних волокон біля кожного базального тіла проходить додаткове мікрофібрилярне сполучне волокно, що з’єднує базальні тіла та кореневу систему джгутика з текою. Ця система отримала назву ризанкіри, тобто кореневого якоря.
Системи мікротубулярних коренів також різноманітні. Наприклад, у пединомонадальної водорості Scourfieldia мікротубулярні корені взагалі відсутні. У родів Pedinomonas та Mantoniella є лише два мікротубулярних корені, у Nephroselmis - три, у Tetraselmis та Pterosperma - чотири. У останнього роду один з коренів складається з багатьох мікротрубочок і пов’язаний з багатошаровою структурою; у представників роду Mesostigma із чотирьох наявних мікротубулярних коренів два складаються з багатьох мікротубочок і з’єднані з багатошаровою структурою (рис. 24.11).
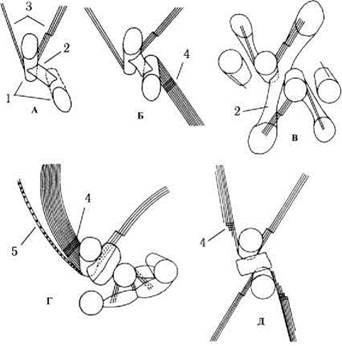
Рис. 24.11. Кореневі системи джгутиків празинофіцієвих водоростей: А - Mantoniella, Б - Nephroselmis, В - Tetraselmis, Г - Pterosperma, Д - Mesostigma. 1 - базальні тіла джгутиків, 2 - дистальне волокно, 3 - мікротубулярні корені, 4 - багатошарова структура, 5 - мікрофібрилярний корінь (за Sym, Pienaar, 1993).
Інші органели. У Prasinophyceae виявлені скоротливі вакуолі, трихоцисти та слизові тільця. Трихоцисти (екструсоми) - це мембранні капсули, що прилягають до плазмалеми і містять згорнуті у вигляді циліндра стрічки. Трихоцисти виявлені у родів Mantoniella, Mamiella, Pyramimonas, Pterosperma, Monomastix.
У Pyramimonas та Halosphaera під плазмалемою знаходяться слизові тільця, які дещо нагадують аналогічні структури рафідофітових водоростей. У деяких видів слизові тільця з’являються лише під час утворення цист. Вони беруть участь у формуванні зовнішнього шару їх оболонки.
Типи морфологічної структури тіла
Переважна більшість празинофіцієвих - це дрібні монадні форми. Проте відомі також гемімонадні (Prasinocladus) та кокоїдні представники (Bathycoccus). Клітини звичайно поодинокі, і не утворюють колоній або ценобіїв. Багатоклітинні форми в межах Prasinophyceae відсутні, хоча цисти деяких родів (Halosphaera) можуть складатися з багатьох клітин, об'єднаних спільною оболонкою, утворюючи т.з. розеткоподібну фікому.
Розмноження та життєві цикли
У празинофіцієвих водоростей відомі нестатеве та статеве розмноження. Життєві цикли представлені переважно цикломорфозами. У одного виду відомий гаплофазний життєвий цикл з зиготичною редукцією та без зміни поколінь.
Нестатеве розмноження одноклітинних монадних форм відбувається поздовжнім поділом надвоє у рухливому стані. У кокоїдних представників (рід Bathycoccus) поділ проходить шляхом схізотомії за допомогою кільцевої борозни. Розмноження зооспорами зустрічається у представників, що мають теку (наприклад, Tetraselmis). Перед утворенням зооспор материнська клітина скидає джгутики, протопласт ділиться, дочірні клітини одягаються власною текою. Таким чином, при розмноженні зооспорами має місце схізогонія.
Статеве розмноження виявлене лише у Nephroselmis olivacea. Статевий процес у цього виду ізогамний, зигота одягається товстою оболонкою і переходить у стан спокою. Після закінчення періоду спокою у зиготі відбувається мейоз, і вона проростає гаплоїдними монадними клітинами.
Спеціальні генетичні дослідження 66 штамів роду Tetraselmis показали, що частина штамів є гетерозиготними. Оскільки згідно законів Менделя, гетерозиготними можуть бути лише еугамні диплоїдні організми, то на підставі цього був зроблений висновок, що у Tetraselmis статевий процес існує (хоча він поки що не спостерігався).
Цикломорфози у празинофіцієвих водоростей. Переважна більшість празинофіцієвих водоростей є агамними, і їх життєві цикли представлені цикломорфозами.
У деяких празинофіцієвих водоростей під час цикломорфозу чергуються монадні та кокоїдні стадії. Наприклад, клітини роду Pycnococcus протягом більшої частини життєвого циклу позбавлені джгутиків і перебувають у кокоїдному стані. Проте за певних умов джгутики відновлюються і водорість короткочасно переходить у монадний стан. У роду Mantoniella цикломорфоз пов’язаний з чергуванням монадного та пальмелевидного станів. В обох станах клітини здатні до схізотомії.
Цикломорфоз у Pterosperma полягає у чергуванні вегетативних монадних стадій та гігантських цист, що отримали спеціальну назву - фікоми. Під час утворення фікоми клітина втрачає джгутики, під плазмалемою з’являються численні слизові тільця, що швидко збільшуються у розмірі і зливаються, формуючи зовнішній шар оболонки цисти. Хлоропласти багаторазово діляться, проте фікома лишається одноядерною. Вона швидко збільшується у розмірі, сягаючи 200-800 мкм у діаметрі, що в 25-100 разів перевищує розмір вегетативних монадних клітин. У фікомі стає помітним внутрішній шар оболонки, а зовнішній шар утворює два великі крилоподібні вирости. Весь процес займає від двох до чотирнадцяти тижнів. Далі відбувається серія поділів ядра та цитоплазми, і утворюється кілька десятків монадних клітин. Зовнішня оболонка цисти розривається, внутрішня ослизнюється, перетворюючись на нетривкий слизовий міхур, з якого звільняються молоді клітини.
У життєвому циклі Tetraselmis чергуються монадний, гемімонадний, пальмелевидний стани та цисти. У гемімонадному стані клітини зовні повторюють морфологію монад, проте позбавлені джгутиків. Така клітина здатна або перетворитися на зооспорангій, або перейти у пальмелевидний стан чи стан цисти. Пальмелевидні клітини звичайно виділяють слиз і утворюють колонії у вигляді простих або розгалужених трубок; як і гемімонадні клітини, вони здатні утворювати зооспори. Цисти Tetraselmis, на відміну від фіком Pterosperma, не збільшуються у розмірі. Під час проростання з однієї цисти виходять 1-4 монадні клітини.
Система класу
Клас поділяється на п’ять порядків. В основу поділу на порядки покладено, в першу чергу, особливості клітинних покривів, цитоскелету, мітозу та загального плану будови клітини (табл. 24.2).
Таблиця 24.2. Основні систематичні ознаки різних порядків Prasinophyceae
2-кето-цукрові кислоти |
Типи клітинних лусочок |
Типи джгутикових лусочок |
Кількість мікротуб. коренів |
Цисти (ц), фікоми (ф) |
Фікопласт |
|
Pedinomonadales |
- |
- |
+, специф. |
0, 2, 4 |
- |
- |
Mamiellales |
+ |
2+3 |
2+3 |
2 |
- |
- |
Pseudoscourfeldiales |
+ |
1+2+(3) |
1+2+(3) |
3+(БШС) |
- |
- |
Pyramimonadales |
+ |
1+2+3 |
1+2+3 |
4+(БШС) |
(ц,ф) |
- |
Chlorodendrales |
+ |
тека з 1+2 |
1+2+3 |
4 |
(ц) |
+ |
Умовні позначення: "+" - наявні; "-" - відсутні; 1, 2, 3 - внутрішні, проміжні та зовнішні лусочки, відповідно; специф. - специфічні; БШС - багатошарова структура; у дужки взято структури, що зустрічаються не у всіх представників даного порядку.
Pedinomonadales об’єднує одно- та дводжгутикові монадні водорості, поверхня клітин яких позбавлена субмікроскопічних лусочок, хоча останні наявні на джгутиках. На відміну від представників інших порядків, до складу джгутикових лусочок у пединомонадальних водоростей не входять 2-кето-цукрові кислоти. Характерними представниками є роди Scourfieldia, Pedinomonas, Resultor, Monomastix.
Види Scourfieldia дводжгутикові і не мають мікротубулярних коренів. У клітин Pedinomonas джгутик один, від його базального тіла відходять два мікротубулярні корені. Одноджгутикова водорість Resultor має чотири мікротубулярних кореня, що відповідають формулі 3-1-2-2. Рід Monomastix характеризується наявністю одного джгутика та базальних трихоцист (рис. 24.12).
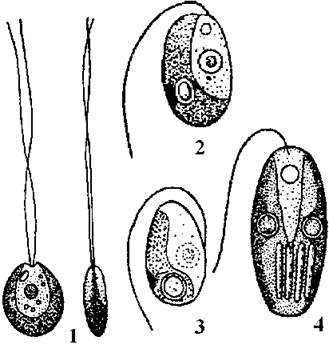
Рис. 24.12. Деякі представники Pedinomonadales: 1 - Scourfieldia, 2 - Pedinomonas, 3 - Resultor, 4 - Monomastix (за Дедусенко-Щеголева и др., 1959; Moestrup, 1991).
На думку деяких дослідників, педіномонадальні водорості рано відокремились від інших празинофіцієвих водоростей і можуть розглядатися як самостійний клас зелених водоростей - Loxophyceae або Pedinophyceae. Молекулярно-філогенетичні реконструкції частково підтверджують цю гіпотезу.
Mamiellales об’єднує водорості, у яких монадні стадії мають два гетероконтні джгутики; у деяких представників коротший джгутик редукований, і від нього зберігається лише базальне тіло. Коренева система складається з двох мікротубулярних коренів. Як клітини, так і джгутики вкриті одним-двома шарами лусочок, причому шар внутрішніх дрібних лусочок відсутній. Цисти не виявлені.
Клітини роду Mamiella несуть два латеральні джутики. Mantoniella має один джгутик, що відходить від невеличкого виступу. Клітини Bathycoccus взагалі позбавлені джгутиків (рис. 24.13).
Мамієллальні водорості за цитологічними ознаками вважаються найбільш примітивною групою зелених водоростей, що дала початок іншим лініям празинофіцієвих.
Pseudoscourfieldiales включає водорості, чиї клітини та джгутики вкриті внутрішніми та проміжними (інколи - також і зовнішніми) лусочками. Монадні стадії мають два гетероконтні джгутики, з базальними тілами яких пов’язані три мікротубулярні корені. У одного з видів є ізогамний статевий процес. Зигота за будовою та хімічним складом оболонки нагадує цисту, хоча останні у представників порядку не виявлені.
У Pseudoscourfieldia джгутики виходять з апікальної заглибини, джгутикові корені мають формулу 4-3-1. У видів роду Nephroselmis джгутики розташовані латерально, формула коренів має вигляд 3-4-8, з третім коренем асоційована структура, подібна до багатошарової (рис. 24.13).
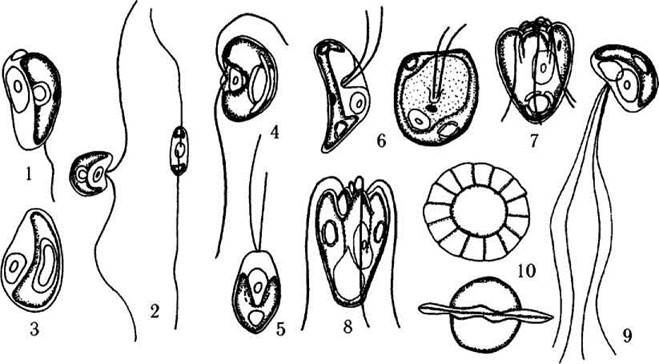
Рис. 24.13. Деякі представники Mamiellales (1-3), Pseudoscourfieldiales (4,5) та Pyramimonadales (6-10): 1 - Mantoniella, 2 - Mamiella, 3 - Bathycoccus, 4 - Nephroselmis, 5 - Pseudoscourfieldia, 6 - Mesostigma, 7 - Pyramimonas, 8 - Halosphaera, 9,10 - Pterosperma (10 - фікома) (за Sym, Pienaar, 1993).
Вважають, що представники Pseudoscourfieldiales могли дати початок зеленим водоростям з асиметричною системою джгутикових коренів, тобто бути пращурами класу Charophyceae.
Pyramimonadales об’єднує водорості з двома, чотирма, зрідка - вісьмома і навіть шістнадцятьма джгутиками, що виходять із джгутикової заглибини. Звичайно у дорослих клітин джгутики ізоконтні, проте у молодих гетероконтні. Коренева система включає чотири мікротубулярні корені, інколи - з багатошаровою структурою та поперечно-смугастим мікрофібрилярним коренем. Клітини пірамімонадальних водоростей завжди вкриті внутрішніми та зовнішніми (часто також і проміжними) лусочками, проте не формують теки. На джгутиках внутрішній шар лусочок є, інші - є або відсутні. У багатьох представників виявлено цисти, у деяких родів - фікоми.
Види роду Mesostigma мають два джгутики, що виходять із латеральної заглибини; джгутикова коренева система хрестоподібна та відповідає формулі 6-4-6-4; з одним із коренів пов’язана багатошарова структура.
Клітини Pyramimonas несуть чотири, вісім або навіть шістнадцять джгутиків, що починаються в апікальній заглибині; коренева система також хрестоподібна, проте позбавлена багатошарової структури.
Дорослі монадні клітини роду Halosphaera мають чотири апікальні джгутики і нагадують види роду Pyramimonas. Коли вегетативні клітини діляться повільно, то дочірні клітини чотириджгутикові, подібні до материнських. Проте у випадку швидкого поділу дочірні клітини зменшуються у розмірах, і мають лише два або навіть один джгутик. Коренева система хрестоподібна, один з коренів утворює багатошарову структуру. Для роду характерні фікоми особливої, розеткоподібної будови.
Види роду Pterosperma також мають чотири джгутики, які, проте розташовуються латерально. Джгутикова коренева система складається з чотирьох хрестоподібно розташованих коренів та п’ятого латерального кореня з багатьох мікротрубочок, що пов’язаний з багатошаровою структурою. Даному роду притаманна здатність утворювати фікоми з крилоподібними виростами. Викопні рештки, що нагадують фікоми такого вигляду знайдено в осадових породах, вік яких складає 1.2 млрд років (рис. 24.13).
Pyramimonadales розглядається як порядок, який, вірогідно, дав початок класам ульвофіцієвих та сифонофіцієвих водоростей. На користь цього припущення свідчить, зокрема, те, що у пірамімонадальних водоростей виявлено ксантофіли сифонеїн та сифоноксантин, а деякі стадії розвитку фікоми мають риси схожості з окремими стадіями онтогенезу дазикладальних водоростей з класу сифонофіцієвих.
Chlorodendrales. До складу порядоку входять монадні та гемімонадні водорості, у яких внутрішні та проміжні лусочки частково зливаються, утворюючи теку. Монадні клітини мають чотири ізоконтні джгутики, що виходять з апікальної заглибини. Коренева система джгутиків хрестоподібна, відповідає формулі 4-2-4-2, багатошарова структура не виявлена. Цитокінез відбувається за участю фікопласта. На відміну від інших порядків, розмноження відбувається зооспорами, причому тільки у нерухомому стані. Цисти відомі, проте фікоми не виявлені.
Монадний тип структури властивий видам родів Scherffelia та Tetraselmis. Відміни між цими родами простежуються за будовою мікрофібрилярних коренів та за наявністю піреноїдів - у першого роду вони відсутні, у другого є (рис. 24.14).
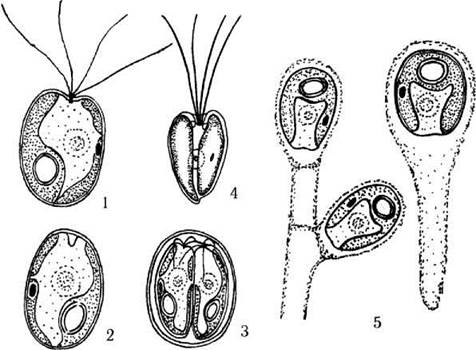
Рис. 24.14. Деякі представники Chlorodendrales: 1-3 - Tetraselmis (1 - монадна клітина, 2 - гемімонадний стан перед початком зооспороутворення, 3 - зооспорангій), 4 - Scherffelia, 5 - Prasinocladus (1-3, 5 - ориг., 4 - за Sym, Pienaar, 1993).
Представники роду Prasinocladus мають гемімонадний тип структури. Клітини знаходяться всередині слизових трубок, і хоча позбавлені джгутиків, проте зберігають базальні тіла та апікальну заглибину. Розмноження відбувається зооспорами, які за зовнішнім виглядом і за ультратонкою будовою практично не відрізняються від монадних клітин Tetraselmis.
Вважається, що хлородендральні водорості могли дати початок класам зелених водоростей з хрестоподібною кореневою системою та цитокінезом, що відбувається за участю фікопласта, тобто Chlorophyceae та Trebouxiophyceae. Це припущення частково підтверджується молекулярно-філогенетичними аналізами за ядерним геном, що кодує 18S субодиницю цитоплазматичних рибосом.
Поширення та екологія
Празинофіцієві водорості мешкають переважно у морях, і лише деякі представники зустрічаються у прісних та гіпергалінних водоймах або в засолених ґрунтах. Морські форми можна знайти у супраліторальних ваннах на океанічних узбережжях. Тут інколи спостерігаються локальні зелені "цвітіння", обумовлені масовим розвитком видів Pyramimonas та Tertaselmis. Відомі випадки, коли празинофіцієві водорості викликали "цвітіння" на досить великих акваторіях - наприклад, у бухтах Фінської затоки (Tetraselmis) або Японського моря (Pyraminonas).
Дрібні монадні та кокоїдні водорості з порядків Mamiellales, Pseudoscourfieldiales та Pyramimonadales є типовими представниками фітопланктону відкритого океану і, як вважають, відіграють помітну роль у загальнопланетарному балансі кисню та вуглецю. Деякі роди (Halosphaera, Pterosperma) у монадному стані мешкають у планктоні, проте у стані фікоми переходять до нейстонного способу життя.
Морські колоніальні форми входять до складу бентосу та перифітону. Так, на узбережжі Чорного моря в обростаннях бурої водорості Cystoseira досить часто можна спостерігати представників роду Prasinocladus.
У гіпергалінних водоймах та засолених ґрунтах інколи масово розвиваються види Pedinomonas, викликаючи зелене "цвітіння" рапи і приймаючи участь в утворенні лікувального мулу.
Прісноводні празинофіцієві водорості з родів Pyramimonas, Nephroselmis та Tetraselmis можна спостерігати в ефемерних водоймах, забруднених органічними речовинами, де вони разом з іншими джгутиковими спричиняють "цвітіння" води.
Крім того, празинофіцієві досить часто зустрічаються як фотосинтезуючі ендосимбіонти тубелярій (Tetraselmis convoluta), радіолярій (деякі мамієляльні водорості), безбарвних динофітових водоростей, зокрема ноктилюки.
1 Детальніший огляд різних варіантів цитологічних систем подано у книзі "Водоросли. Справочник" (Вассер, Кондратьева, Масюк и др., 1989).