Альгологія - Костіков І.Ю. - 2009-2013
Розділ 24. Зелені водорості – Chlorophyta
24.2. Хлорофітна лінія
24.2.2. Клас Trebouxiophyceae
Цей клас встановлений порівняно недавно - у 80-х роках. Саме тоді на основі даних електронної мікроскопії було зроблено припущення про існування у межах класу Chlorophyceae самостійної еволюційної лінії, що спочатку отримала назву порядку Pleurastrales, далі - класу Pleurastrophyceae, і нарешті - класу Trebouxiophyceae. Сьогодні цей клас нараховує біля 250 видів кокоїдних, сарциноїдних, нитчастих та гетеротрихальних водоростей, що мешкають переважно у позаводних екотопах - у ґрунтах, в аерофітоні, входять до складу лишайників, рідше зустрічаються як симбіонти прісноводних безхребетних тварин (найчастіше - інфузорій та губок). Клас представляє лінію зелених водоростей, в межах якої цитокінез відбувається за участю фікопласту, базальні тіла джгутиків зміщені проти годинникової стрілки та мають орієнтацію 11-5, джгутикова коренева система хрестоподібна, а монадні клітини ніколи не несуть субмікроскопічних лусочок.
Ознаки та характеристика класу
Біохімічні особливості
Склад пігментів та продуктів асиміляції такий самий, як і у представників Chlorophyceae, і таким чином, специфічні біохімічні особливості в межах класу відсутні.
Цитологічні особливості
Клітинні покриви. Клітини требуксієфіцієвих завжди вкриті целюлозно- пектиновою оболонкою, яка, звичайно, має тришарову будову і у багатьох представників (зокрема, у Chlorella) містить додатковий шар спорополленіну. Монадні клітини, представлені лише зооспорами та зрідка - гаметами, завжди вкриті лише плазмалемою, після зупинки набувають сферичної форми і відносяться до п-типу.
Субмікроскопічні лусочки як на клітинах, так і на джгутиках у требуксієфіцієвих водоростей відсутні.
Ядерний апарат та особливості мітозу. Мітоз у межах класу напіввідкритий (у більшості видів) або закритий (у празіолальних водоростей), веретено поділу у телофазі звичайно швидко руйнується (вийнятком є празіолальні водорості, у яких телофазне веретено є персистентним). Центриолі у требуксієфіцієвих водоростей наявні лише у зооспороутворюючих видів; у автоспорових представників вони відсутні. Якщо центриолі є, вони не приймають безпосередньої участі в утворенні веретена поділу, проте відіграють суттєву роль у побудові фікопласту.
Цитокінез відбувається за участю борозни поділу, яка завжди починає формуватися лише з одного боку клітини. У кокоїдних та сарциноїдних форм по закінченні цитокінезу дочірні протопласти ще у материнській клітині виробляють власні оболонки, і таким чином, поділ відповідає схізогонії. У нитчастих та гетеротрихальних видів вегетативний поділ являє собою справжню цитотомію (рис. 24.32).
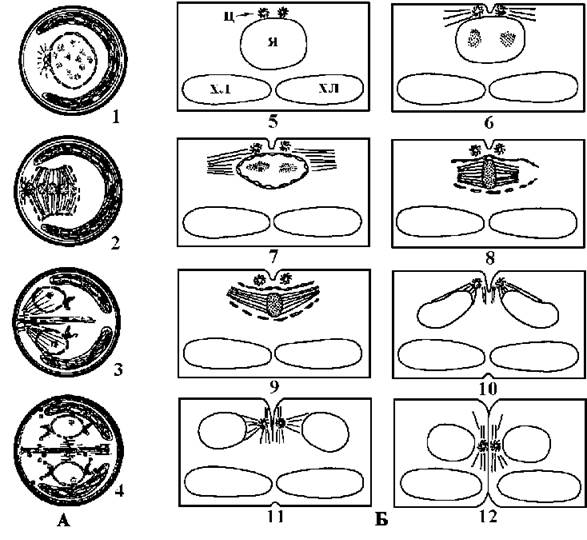
Рис. 24.32. Поділ клітин у сарциноїдної водорості Friedmannia israeliensis (A) та гетеротрихальної водорості Leptosira obovata (Б): 1, 5-7 - профаза; 2, 8, 9 - метафаза; 3, 10, 11 - телофаза; 4, 12 - рання інтерфаза; я - ядро, хл - хлоропласт, ц - центріолі (за Lokhorst, Segaar, Star, 1989; Molnar, Stewart, Mattox, 1975).
Джгутиковий апарат. Монадні клітини представлені зооспорами та зрідка - гаметами, і мають 2 (зрідка - 4) ізоконтних та ізоморфних джгутика. Звичайно монадні клітини дорзовентральні та сплощені у поздовжньому напрямку.
Перехідна зона джгутиків містить зірчасту структуру, спіралеподібні стрічки та циліндри у перехідній зоні відсутні. Базальні тіла відносно короткі, зміщені проти годинникової стрілки (11-5 конфігурація), і досить помітно перекриваються одне з одним. Верхня перетинка, що з’єднує базальні тіла, поперечно-смугаста (рис. 24.33).
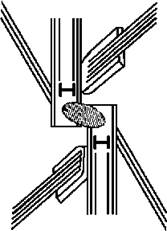
Рис. 24.33. Схема будови кореневої системи джгутиків требуксіофіцієвих водоростей (згідно до мікрофото Deason, Floyd, 1987).
Коренева система хрестоподібна і звичайно відповідає формулі 4-2-4-2 або 5-2-5-2. На верхівках товстих коренів виявлено т.з. колончасту структуру, яка дещо нагадує багатошарову структуру празинофіцієвих та харофіцієвих. Ризопласти достовірно не виявлені, хоча у деяких представників знайдено структуру, що відповідає рудиментарному поперечно-смугастому мікрофібрилярному кореню.
Типи морфологічних структур
В межах класу представлено водорості з чотирма типами морфологічних структур: кокоїдним, сарциноїдним, нитчастим та гетеротрихальним.
Кокоїдні водорості є переважаючою групою. Їх клітини звичайно сферичні, поодинокі, колоній та ценобіїв звичайно не утворюють. Сарциноїдні водорості нечисленні, звичайно мають таломи у вигляді пакетів (Pleurastrosarcina) або тетрад (Friedmannia).
Нитчасті водорості досить численні, як правило, утворюють короткі нитки, що легко розпадаються на окремі клітини (наприклад, Stichococcus, Diplosphaera), або довгі нитки з тенденцією переходу до дво- або багаторядності (Prasiola, Schizogonium). Гетеротрихальні форми мають добре розвинену систему розгалужених висхідних ниток, і дуже редуковані сланки нитки, які звичайно представлені лише однією базальною клітиною (Leptosira, Microthamnion).
Розмноження та життєві цикли
Для кокоїдних представників основним способом розмноження є нестатеве розмноження за допомогою апланоспор та рідше - зооспор. Нитчасті та гетеротрихальні форми розмножуються переважно вегетативно - за рахунок вегетативного поділу з подальшою фрагментацією ниток, і лише за виняткових умов - зоо- та апланоспорами. У деяких празіолальних водоростей є статеве розмноження, що являє собою оогамію (Prasiola). Водорості з сарциноїдним типом структури розмножуються як за рахунок фрагментації багатоклітинних агрегатів (пакетів, тетрад та комплексів з них), так і зооспорами та апланоспорами.
Переважаючим типом життєвого циклу є простий цикломорфоз. У видів роду Prasiola, що мають статевий процес, життєвий цикл є гаплодиплофазним, з ізоморфною зміною поколінь та соматичною редукцією.
Система класу
Оскільки клас встановлено порівняно недавно, і переважно - на підставі цитологічних та молекулярно-біологічних критеріїв, система класу розроблена поки що недосконало. Сьогодні в межах класу за результатами аналізу ядерного гену, що кодує 5S та 18S субодиниці рибосомальної РНК, встановлено до 10 незалежних ліній розвитку, що вірогідно відповідають таксонам рангу порядку. Проте лише чотири лінії отримали статус порядків, дві лінії мають робочі назви, які проте не відповідають вимогам Міжнародного Кодексу Ботанічної номенклатури (зокрема, не мають латинських діагнозів та типу). Інші лінії розглядаються як групи нез’ясованого таксономічного положення.
Систематичними критеріями для розмежування встановлених порядків за фенотипічними ознаками є, в першу чергу, здатність до розмноження за допомогою зооспор, тип морфологічної структури, наявність гетерополярності клітин або таломів, пов’язаної зі здатністю до прикріпленого способу життя, тип мітозу, наявність чи відсутність персистентного телофазного веретена (табл. 24.4).
Таблиця 24.4. Основні систематичні ознаки порядків требуксієфіцієвих водоростей
Ознака → Порядок ↓ |
Тип структури |
Гетерополярність |
Зооспори |
Мітоз |
Телофазне веретено |
Chlorellales |
кокоїдний |
- |
- |
НВ |
- |
Trebouxiales |
кокоїдний, сарциноїдний |
- |
+ |
НВ |
- |
*Stichococcales |
нитчастий |
- |
- |
НВ |
- |
*Leptosirales |
гетеротрихальний |
- |
+ |
НВ |
- |
Microthamniales |
кокоїдний, нитчастий |
+ |
+ |
НВ |
- |
Prasiolales |
нитчастий |
- |
- |
З |
+ |
Примітки: НВ - напіввідкритий мітоз, З - закритий мітоз. Знаком (*) позначено таксони, які отримали назви без урахування вимог МКБН, і тому не є таксономічно законними.
Chlorellales. Порядок об’єднує кокоїдні, переважно - сферичні автоспорові водорості з напіввідкритим мітозом та зникаючим у телофазі веретеном. Клітини хлореллальних водоростей не є гетерополярними.
Звичайно види Chlorellales мають дуже дрібні розміри (до 8-10 мкм), розмножуються за допомогою 2-8 автоспор, що звільняються після розриву оболонки спорангію. Серед представників порядку відомі як фотоавтотрофні (Chlorella), так і облігатно гетеротрофні (Prototheca) форми.
Клітини Chlorella у дорослому стані сферичні, містять одне ядро, парієтальний хлоропласт з піреноїдом (Ch.vulgaris) або без нього (Ch.minutissima) (рис. 24.33).
Види роду Chlorella є характерним компонентом позаводних ценозів. Їх можна знайти у ґрунтах, на корі дерев, рідше - водних біоценозах, звичайно забруднених органічною речовиною. В межах роду спостерігається тенденція переходу до симбіотичного способу життя: зокрема, деякі Chlorella живуть у клітинах інфузорій, губок, у таломах лишайників. У багатьох країнах види Chlorellaвведені у промислову культуру і вирощуються як сировина для виготовлення вітамінних домішок або дієтичної їжі.
Prototheca є безбарвною гетеротрофною водорістю, морфологічно подібною до Chlorella. Розмножується вона за допомогою 2-8 автоспор. Прототека звичайно паразитує на ссавцях (в тому числі - на людини), і викликає характерні захворювання шкіри - прототекози. У вільному стані її можна знайти у позаводних ценозах - на корі дерев, вологих стінах, зрідка - у ґрунті, якщо субстрат містить достатню кількість розчиненої органічної речовини.
Trebouxiales. Порядок об’єднує кокоїдні та сарциноїдні водорості, що за систематичними ознаками подібні до Chlorellales, проте здатні до розмноження за допомогою зооспор. Всі требуксіальні водорості зустрічаються лише у позаводних ценозах. Основними родами порядку є Trebouxia, Myrmecia та Friedmannia (рис. 24.34).
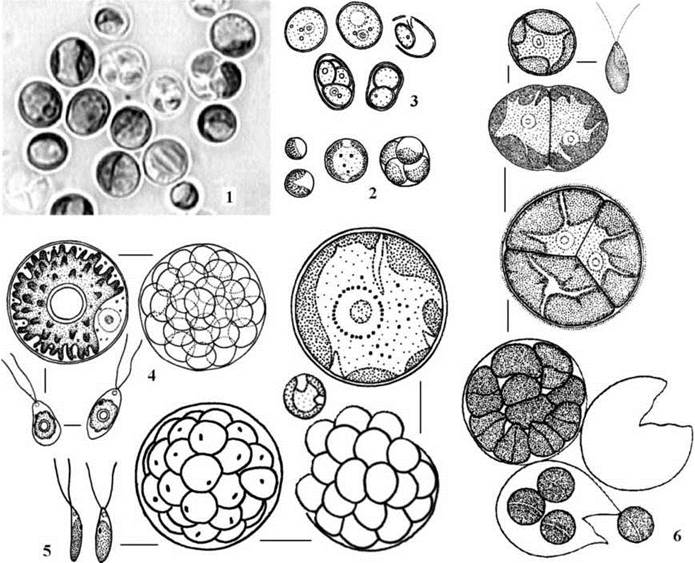
Рис. 24.34. Деякі характерні представники Chlorellales (1-3) та Trebouxiales (4-6): 1 - Chlorella vulagaris; 2 - Chlorella minutissima; 3 - Prototheca sp. ; 4 - Trebouxia crenulata; 5 - Myrmecia macronucleata; 6 - Friedmannia israeliensis (1-3, 5 - Костіков, ориг.; 4 - Михайлюк, 2000; 6 - за Chantanachat, Bold, 1962).
Trebouxia має сферичні клітини з центральним зірчастим або осьовим хлоропластом, піреноїдом та одним ядром. Розмноження відбувається голими дводжгутиковими зооспорами та апланоспорами. Рід містить біля 50 видів, що є найтиповішими фікобіонтами лишайників.
Myrmecia за морфологією та розмноженням нагадує попередній рід, проте має пристінний лопатевий хлоропласт позбавленй піреноїду. Види цього роду зустрічаються і як фікобіонти лишайників, і у вільному стані - в ґрунтах та на кам’янистих субстратах в аерофітоні.
Особливістю Friedmannia є її здатність утворювати тетради клітин та комплекси, що складаються з тетрад, тобто ця водорість має сарциноїдний тип структури. Будова клітин у Friedmannia така ж сама як і у Myrmecia. Розмноження відбувається за допомогою голих дводжгутикових зооспор, апланоспор або вегетативно - внаслідок розпаду тетрад на поодинокі клітини.
Stichococcales. До порядку входять водорості, клітини яких здатні до поділу в одній або двох площинах з утворенням коротких ниток. Всі відомі представники порядку є поширеними аерофітними та ґрунтовими водоростями.
Водорості роду Stichococcus утворюють короткі однорядні нитки, що легко розпадаються на поодинокі одноядерні клітини. Кожна клітина містить пристінний дископодібний хлоропласт позбавлений піреноїду; на полюсах клітин звичайно є по одній краплині олії. Розмноження виключно вегетативне, нестатеве та статеве розмноження не відомі. Види цього роду найчастіше зустрічаються на корі дерев, на поверхні плодових тіл старих трутовиків, і досить часто спричинюють їх "цвітіння". Рідше Stichococcus знаходять у ґрунтах та на кам’янистих субстратах в аерофітоні (рис. 24.35).
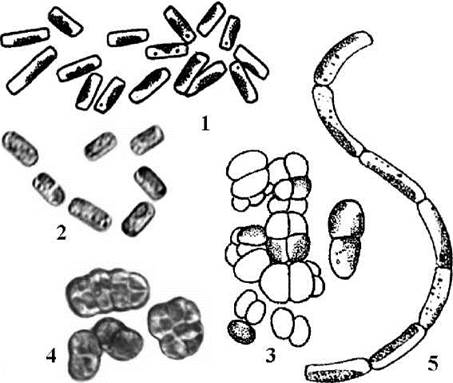
Рис. 24.35. Деякі характерні представники Stichococcales: 1, 2 - Stichococcus bacillaris; 3, 4 - Diplosphaera chodatii; 5 - Gloeotila spiralis (1, 5 - за Chodat, 1902, 1913; 3 - за Vischer, 1945; 2,4 - Костіков, ориг.).
Види роду Gloeotila, хоча і нагадують Stichococcus за наявністю циліндричних клітин з пристінним хлоропластом, позбавленим піреноїду, проте на відміну від останнього утворюють довгі нитки, що звичайно не розпадаються на поодинокі клітини. Цікаво, що за молекулярними даними, з приблизно 15 видів, описаних в цьому роді, до Trebouxiophyceae належать лише ті види, які не мають зооспор і мешкають у позаводних екотопах.
Клітини роду Diplosphaera здатні до поділу у двох площинах. Внаслідок цього утворюються не тільки однорядні нитки, але й пакетоподібні тримірні комплекси.
Leptosirales. Включає водорості з гетеротрихальним типом структури, що розмножуються як вегетативно, так і за допомогою зооспор п-типу та апланоспор. Як і представники попередніх порядків, лептозиральні водорості мешкають переважно у позаводних біотопах.
Види роду Leptosira мають таломи, що складаються з коротких висхідних ниток та розвиненої системи сланких ниток, що досить часто утворюють паренхіматозні платівки (наприклад, L.terrestris, L.obovata). У водних культурах утворюються численні дводжгутикові зооспори, що при проростанні прикріплюються до поверхневої плівки води і проростають у короткі дво- або триклітинні нитки, які здатні тривалий час вести нейстонний спосіб життя. Подальший розвиток ниток у багатоклітинні таломи відбувається тоді, коли клітини осідають на субстрат вище урізу води, тобто переходять до аерофітного способу життя.
В роду Leptosira описано також чимало облігатно прісноводних перифітонних видів, проте всі ці види у цитологічному та молекулярно-біологічному відношенні лишаються не дослідженими і їх належність до Trebouxiophyceae не є остаточно доведеною.
Існує думка, щодо групи лептозіроподібних требуксієфіцієвих водоростей після відповідних молекулярних та електронно-мікроскопічних досліджень увійде також багато інших аерофітних гетеротрихальних водоростей (наприклад, збудники аерофітного "цвітінні" кори дерев та кам’янистих субстратів з родів Desmococcus, Apatococcus, Pseudopleurococcus та інші), які за класичною системою входять до Chlorophyceae, проте за сучасною системою розглядаються як таксони невизначеного систематичного положення (рис. 24.36).
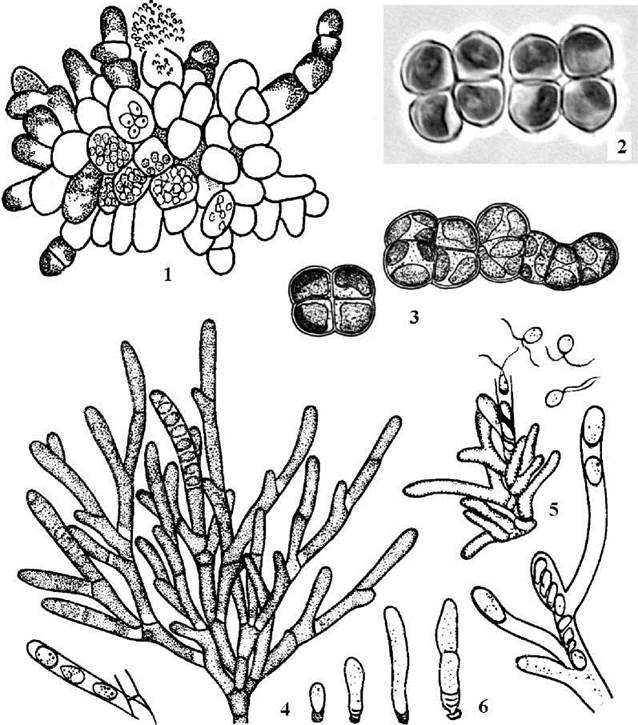
Рис. 24.36. Деякі представники Leptosirales (1-3) та Microthamniales (4-6): 1 - Leptosira obovata; 2 - Apatococcus lobatus; 3 - Desmococcus olivaceus; 4-6 - Microthamnion kuetzingianum (4 - дорослий талом, 5 - вихід зооспор, 6 - початкові стадії розвитку талому) (1, 4-6 - за Мошкова, 1979; 2 - Костіков, ориг.; 3 - за Коршиков, 1953).
Microthamniales. До порядку входять кокоїдні та розгалужені нитчасті водорості, що здатні розвиватися як у водномих, так і в наземних умовах, тобто є амфібіальними організмами.
Специфічною особливістю представників порядку є полярність їх таломів. Наприклад, у кокоїдної водорості Fusochloris perforata, що мешкає звичайно на снігу, клітини диференційовані на округлу верхівкову та витягнуту та звужену базальну частину.
У гетеротрихальних водоростей роду Microthamnion талом складається з базальної клітини, що утворює прикріплювальну підошву і дещо нагадує клітини Fusochloris, та системи розгалужених висхідних ниток. Кожна клітина має один пристінний хлоропласт без піреноїду і є одноядерною (рис. 24.36).
Обидва роди розмножуються переважно нестатевим шляхом - за допомогою голих дводжгутикових зооспор.
Prasiolales. Порядок об’єднує нитчасті водорості, у яких простежується тенденція до утворення багаторядних ниток внаслідок поділу клітин не тільки у поперечному, але й поздовжньому напрямку. Клітини мають центральний хлоропласт з погано помітним піреноїдом. Розмноження відбувається фрагментацією таломів або апланоспорами. У деяких видів виявлено статевий процес оогамного типу. Систематичною ознакою, що чітко відрізняє празиоляльні водорості від інших груп Trebouxiophyceae є наявність закритого мітозу та тривале збереження веретена поділу у телофазі.
Види з Prasiolales зустрічаються у позаводних ценозах, звичайно на субстратах, багатих на амонійні сполуки та солі калію - на посліді на пташиних базарах, на ґрунті, забрудненому органічною речовиною та ін.
Для роду Schizogonium характерні переважно однорядні нитки; лише на деяких ділянках талому клітини здатні до поздовжнього поділу і утворюють короткі дво- або трирядні сегменти (рис. 24.37).
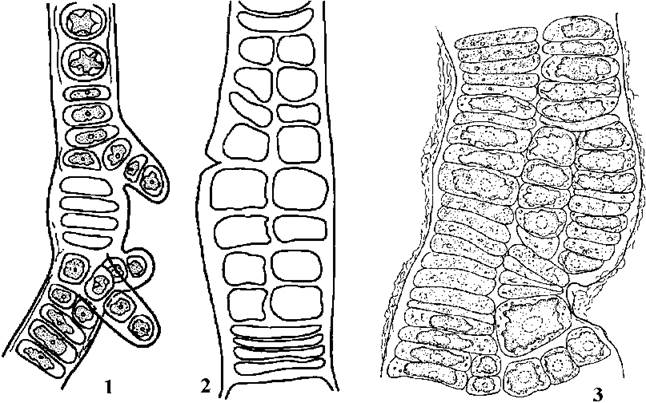
Рис. 24.37. Характерні представники Prasiolales. 1, 2 - Schizogonium murale; 3 - Prasiola velutina (1, 2 - Масюк, Костиков, 1984; 5 - за Lokhorst, Starr, 1988).
У видів роду Prasiola, навпаки, більшість клітин здатна до поділу у двох площинах, внаслідок чого утворюються багаторядні таломи пластинчастої форми. У деяких видів відоме статеве розмноження.
Так, у P.stipitata вегетативна слань диплоїдна. При підготовці до статевого розмноження група крайових клітин слані редукційно ділиться, далі відбувається серія мітозів, внаслідок чого на диплоїдному таломі утворюються велика за площею зона гаплоїдних вегетативних клітин, тобто редукційний поділ є соматичним. Одна частина гаплоїдних клітин надалі диференціюється в антеридії, що дають початок дводжгутиковим сперматозоїдам; друга частина розвивається в оогонії, в яких утворюються нерухомі сферичні яйцеклітини. Після дозрівання при наявності рідкої вологи сперматозоїди та яйцеклітини виходять у зовнішнє середовище, де і відбувається копуляція (рис. 24.38).
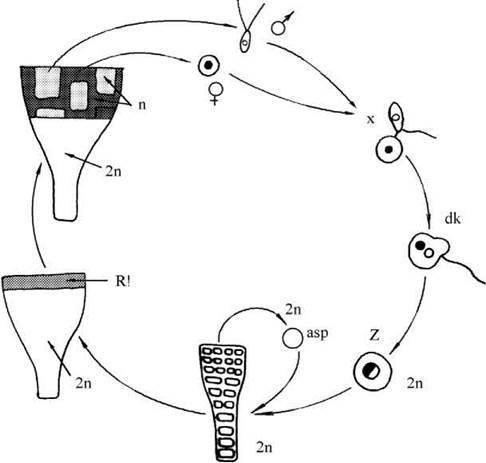
Рис. 24.38. Життєвий цикл Prasiola stipitata. Asp - апланоспора; R! - редукційний поділ; n, 2n - гаплоїдні та диплоїдні клітини, відповідно; dk - дикаріонтична планзигота; Z - зріла зигота (за Friedmann, 1959, 1964, 1969).
Процес копуляції у Prasiola унікальний - сперматозоїд підпливає до яйцеклітини, поникає в неї одним джгутиком, далі одним боком зливається з яйцеклітиною. При цьому каріогамія одразу не відбувається, зигота лишається двоядерною. У зиготи зберігається один джгутик - той, що не проник у яйцеклітину. Протягом кількох діб за допомогою цього джгутику зигота здатна до повільного руху. Після зупинки ядра у зиготі одразу зливаються, і вона без періоду спокою починає проростати у новий диплоїдний талом.