Альгологія - Костіков І.Ю. - 2009-2013
Розділ 24. Зелені водорості – Chlorophyta
24.2. Хлорофітна лінія
24.2.3. Клас Ulvophyceae
Клас об’єднує переважно морські бентосні та перифітонні водорості, хоча в межах класу представлені також і прісноводні та аерофітні форми. Майже всі ульвофіцієві водорості мають багатоклітинні таломи, переважаючі типи структури - нитчастий, гетеротрихальний та сифонокладальний. Ulvophyceae представляє лінію еволюції зелених водоростей, у якої цитокінез відбувається за типом кільцевої борозни, коренева система джгутиків хрестоподібна, а базальні тіла зміщені проти годинникової стрілки.
До класу входить біля 80 родів, що об’єднують більш ніж 1000 видів.
Ознаки та характеристика класу
Біохімічні особливості
Специфічною особливістю є наявність у багатьох морських сифонокладальних представників додаткового специфічного ксантофілу - сифоноксантину. Цей ксантофіл зустрічають також у більшості видів з класу сифонофіцієвих та у деяких празинофіцієвих водоростей.
Цитологічні особливості
Клітинні покриви. Клітини завжди вкриті целюлозно-пектиновою або целюлозною оболонкою. Монадні клітини представників найбільш примітивного порядку - Codiolales - на плазмалемі мають також субмікроскопічні органічні лусочки, що за походженням та морфологією ідентичні внутрішнім лусочкам празинофіцієвих. Монадні стадії представників інших порядків голі, належать до п-типу.
Ядерний апарат та особливості мітозу. Мітоз у межах класу закритий, веретено утворюється за участю центріолей і зберігається тривалий час у телофазі. У ульвофіцієвих відомо два варіанти поведінки хроматину у метафазі: у першому варіанті хроматин конденсується у справжні хромосоми, що вистроюються в екваторіальній площині і далі розходяться за типовою для еукаріот схемою; при другому варіанті хроматин конденсується в одну щільну платівку з кількома каналами, через які проходять пучки мікротрубочок інтерзонального веретена. Частина мікротрубочок прикріплюється до кінетохорів платівки, і зумовлює подальший розподіл хроматину між дочірніми ядрами (рис. 24.39).
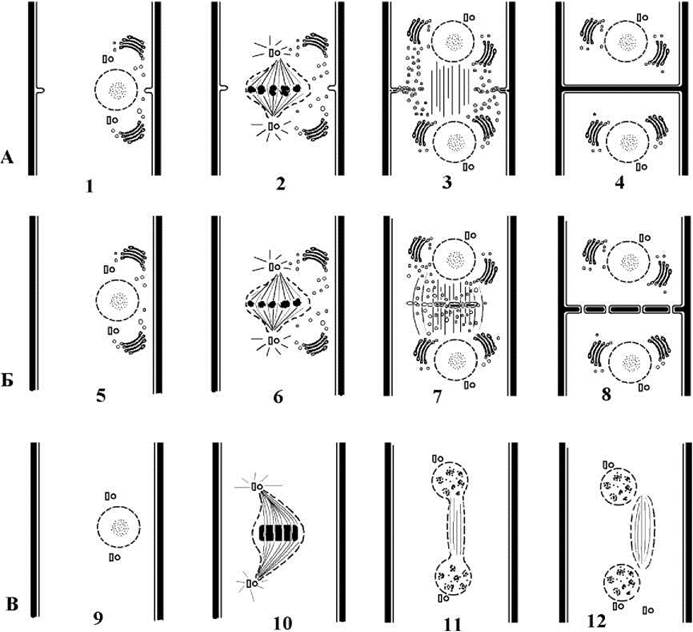
Рис. 24.39. Різноманітність мітозу та цитокінезу у ульвофіцієвих водоростей. А - кодіоляльні, акросифональні та ульвальні водорості: хромосоми у метафазі помітні, цитокінез відбувається шляхом вростання борозни поділу без участі фіко- або фрагмопласту; Б - трентеполіальні водорості: хромосоми у метафазі помітні, цитокінез відбувається за участю фрагмопласту шляхом утворення клітинної платівки, поперечна перегородка має пори з плазмадесмами; В - кладофоральні водорості: хроматин конденсується у щільну платівку, у якій окремі хромосоми не помітні, в анафазі та телофазі ядро сильно витягується в довжину; по закінченні мітозу веретено відокремлюється від дочірніх ядер і деякий час оточено залишками оболонки материнського ядра. 1, 5, 9 - рання профаза, 2, 6, 10 - метафаза, 3, 7, 11 - телофаза, 4, 8, 12 - рання інтерфаза (схематизовано згідно до Hori, Enomoto, 1978; Sluiman et al., 1980; Chapman, Henk, 1986).
Цитокінез відбувається за допомогою кільцевої борозни або клітинної платівки. Ріст як кільцевої борозни, так і клітинної платівки зумовлений, перш за все, діяльністю комплексу Гольджі: пухирці комплексу Гольджі з матеріалом майбутньої клітинної перегородки транспортуються до площини поділу, і там зливаються або з зачатковою кільцевою борозною, обумовлюючи її ріст, або між собою, утворюючи клітинну платівку. При цитокінезі за участю кільцевої борозни пухирці комплексу Гольджі транспортуються без участі фікопласту та фрагмопласту, клітинна перегородка вростає центрипетально і позбавлена пор. При цитокінезі за типом клітинної платівки напрямок руху пухирців визначає фрагмопласт, а поперечна перегородка має пори з плазмадесмами.
Джгутиковий апарат. Монадні клітини представлені зооспорами та гаметами, і мають 2 або 4 ізоконтних та ізоморфних джгутика.
Джгутики гладкі, на поперечному перерізі звичайно циліндричні, проте у трентеполіальних водоростей кожний джгутик має латеральний крилоподібний вирост. Перехідна зона містить зірчасту структуру.
Базальні тіла джгутиків видовжені, змішені проти годинникової стрілки (11-5 конфігурація), звичайно сильно перекриваються одне з одним. З латеральної позиції вони розгорнуті майже на 180°. Верхня перетинка, що з’єднує базальні тіла, є гладкою (винятком є кладофоральні водорості).
Коренева система хрестоподібна, кількість мікротрубочок у коренях відповідає формулі 3-2-3-2 або 6-4-6-4. Верхівки коренів, що прилягають до базальних тіл, вкриті тонкими термінальними ковпачками. Крім того, у трентеполіальних водоростей виявлено колончасту структуру. Ця структура подібна до такої ж у требуксієфіцієвих, проте асоціюється не з товстими, а з верхівками тонких коренів (рис. 24.40).
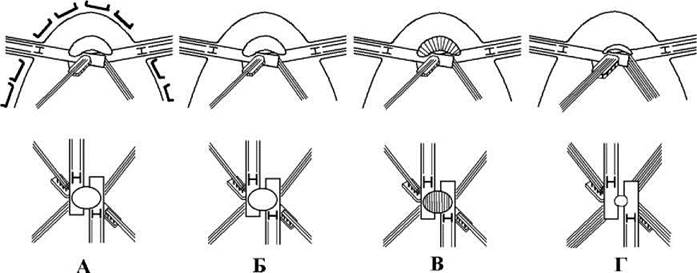
Рис. 24.40. Джгутиковий апарат ульвофіцієвих водоростей при вигляді збоку та зверху: А - Codiolales: монадні клітини вкриті субмікроскопічними лусочками, верхнє з’єднувальне волокно гладке; Б - Ulvales та Acrosiphonales: коренева система подібна до ульвальних, проте субмікроскопічні лусочки відсутні; В - Cladophorales: верхнє з’єднувальне волокно поперечно-смугасте; Г - Trentepohliales: кількість мікротрубочок у джгутикових коренях подвоєна, верхнє з’єднувальне волокно редуковане (згідно до O'Kelly, Floyd, 1984; O'Kelly et al., 1984; Bakker, Lokhorst, 1985; Roberts, 1984).
Таким чином, за абсолютною конфігурацією 11-5 ульвофіцієві нагадують требуксієфіцієві водорості. Проте видовженість, сильне перекривання, майже повна розгорнутість базальних тіл, наявність гладкої верхньої перетинки, наявність у товстих коренях мікротрубочок у кількості кратній трьом, асоційованість колончастої структури з тонкими коренями добре відрізняє джгутикові стадії ульвофіцієвих від требуксієфіцієвих.
Типи морфологічних структур
В межах класу представлено водорості з пятьма типами морфологічних структур: кокоїдним, нитчастим, гетеротрихальним, паренхіматозним та сифонокладальним.
Кокоїдні водорості нечисленні і представлені переважно ендофітними (Chlorocystis) та паразитичними (Chlorochytrium) видами. У деяких паразитичних представників спостерігається тенденція переходу до сифональної будови.
Нитчасті водорості є однією з переважаючих груп у межах класу. Вони звичайно мають однорядні нерозгалужені (Ulothrix) таломи, для деяких форм характерні таломи у вигляді одношарової платівки (Monostroma).
Гетеротрихальні форми досить численні, і представлені в першу чергу двома морфологічними групами: по-перше, водоростями з розвиненими сланкими та висхідними нитками (Spongomorpha); на останніх інколи наявні одноклітинні волоски (хети) з бульбоподібно розширеною основою (Acrochaete), по-друге, з редукованою системою сланких та розвиненою системою висхідних ниток (Trentepohlia). Висхідні нитки у деяких форм можуть утворювати паренхіматозні диски (Cephaleuros).
Водорості зі справжньою паренхіматозною будовою порівняно нечисленні і мають таломи у вигляді двошарових платівок (Ulva) або трубок з внутрішньою порожниною (Enteromorpha).
Сифонокладальні форми представлені досить широко. Таломи можуть мати вигляд нерозгалужених ниток (Rhizoclonium), кущика, утвореного розгалуженими нитками (Cladophora), системи з’єднаних між собою міхурів (Valonia), трубок однакових чи різних розмірів (Siphonocladus) та ін.
Розмноження та життєві цикли
В межах класу широко представлено як нестатеве, так і статеве розмноження. Нестатеве розмноження відбувається фрагментацією таломів, чотириджгутиковими (зрідка - дводжгутиковими) зооспорами, апланоспорами. Відомі також акінети. Статевий процес переважно ізогамний або гетерогамний.
Життєві цикли гаплодиплофазні, з ізо- або гетероморфною зміною поколінь. У багатьох представників зигота проростає в особливий одноклітинний спорофіт, що отримав назву стадії "Codiolum". Досить широко представлені варіанти партеногенетичного розвитку.
Система класу
В межах класу за результатами аналізу різних ядерних та хлоропластних генів встановлено 5 основних груп видів, що можуть розглядатися як таксони рангу порядку (табл. 24.5). Цікаво, що молекулярна (генотипчна) відокремленість цих груп добре корелює з цілим рядом фенотипічних ознак, пов’язаних з ультратонкою будовою клітин (в першу чергу - зооспор та гамет), життєвими циклами та біологією розмноження, певною мірою - морфологією та екологічними особливостями. Вважають, що дивергенція у межах класу відбувалася саме у цих напрямках.
Табл. 24.5. Деякі основні систематичні ознаки порядків ульвофіцієвих водоростей
Ознака → Порядок ↓ |
Тип структури |
Codiolum-стадія |
Зміна поколінь |
СМЛ |
Екологічна група |
Codiolales |
кок., нитч., г-трих. |
+ |
ГМ |
+ |
морські (прісноводні) |
Acrosiphonales |
+ |
ГМ |
- |
морські |
|
Ulvales |
г-трих., пар. |
- |
ІМ |
- |
морські (прісноводні) |
Cladophorales |
- |
ІМ |
- |
морські, прісноводні |
|
Trentepohliales |
- |
ІМ |
- |
Скорочення: ЖЦ - життєвий цикл, СМЛ - субмікроскопічні лусочки, кок. - кокоїдний, нитч. - нитчастий, г-трих. - гетеротрихальний, пар. - паренхіматозний, сиф-клад. - сифонокладальний, IM - ізоморфна зміна поколінь, ГМ - гетероморфна зміна поколінь.
Codiolales (Ulotrichales). Порядок об’єднує біля 90 видів водоростей, що мешкають переважно у морях, хоча окремі представники досить часто зустрічаються і у прісних континентальних водоймах.
Кодіоляльні водорості переважно мають вигляд нерозгалужених ниток або рясно розгалужених кущиків, рідше - поодиноких одно- або багатоядерних клітин чи одношарових платівок. Специфічними ознаками порядку вважаються, в першу чергу, наявність субмікроскопічних лусочок на монадних клітинах та гаплодиплофазного життєвого циклу з гетероморфною зміною поколінь, причому спорофіт представлений диплоїдною стадією Codiolum. Клітинні оболонки містять велику кількість пектинових речовин, і звичайно злегка ослизнюються.
Прикладом кокоїдної одноядерної водорості є морський рід Chlorocystis. Гаметоспорофіт хлороцистісу мешкає у слизових трубках діатомових водоростей або в таломах водоростей-макрофітів; він має сферичні одноядерні клітини з пристінним пальчасто розсіченим хлоропластом, у якому міститься піреноїд (рис. 24.41).
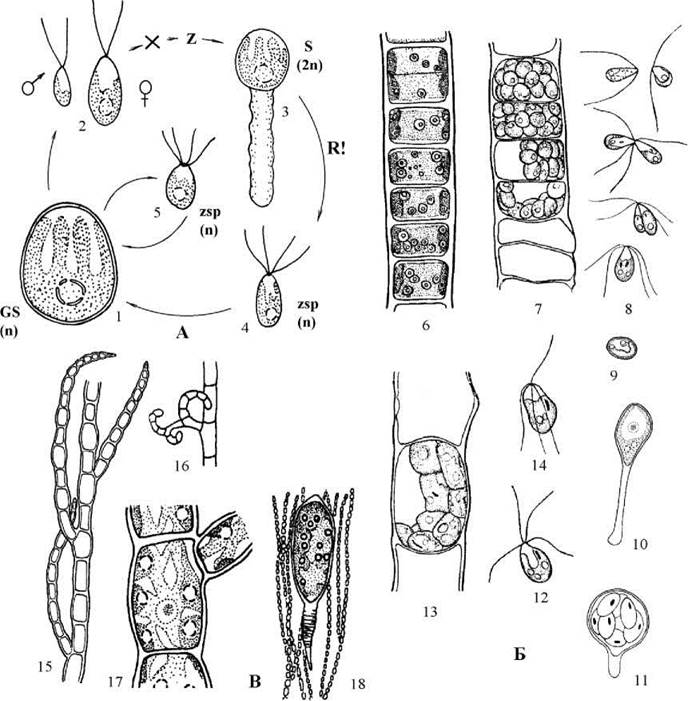
Рис. 24.41. Деякі представники Codiolales: А - життєвий цикл Chlorocystis (1 - гаметоспорофіт; 2 - гамети; 3 - спорофіт, стадія Codiolum; 4 - зооспора спорофіту, 5 - зооспора гаметоспорофіту); Б - Ulothrix (6 - гаметоспорофіт; 7 - утворення гамет; 8 - послідовні стадії копуляції гамет; 9 - зигота; 10 - спорофіт, стадія Codiolum; 11 - утворення зооспор спорофітом; 12 - зооспора спорофтіу; 13 - утворення зооспор гаметоспорофітом; 14 - зооспора гаметоспорофіту); В - Spongomorpha (16 - фрагмент талому гаметофіту; 17 - гачкоподібна бічна гілка - цироїд; 18 - клітина; 19 - багатоядерний спорофіт "Chlorochytrium") (1-5 - за Kornmann, Sahling, 1983; 6-16 - за Мошкова, 1979; 17 - Костіков, ориг., 18 - за Kornmann, 1964).
Нестатеве розмноження відбувається за допомогою 8-32 чотириджгутикових зооспор, які після зупинки проростають у новий гаметоспорофіт. При несприятливих умовах починається статевий гетерогамного типу. Як чоловічі, так і жіночі гамети дводжгутикові, і утворюються у різних клітинах, тобто водорість є роздільностатевою. Після копуляції утворюється зигота з гладкою оболонкою, яка починає проростати у диплоїдний спорофіт - стадію Codiolum: на одному боці формується булавоподібний виріст, в який переходить більша частина протопласту зиготи. Клітина росте, помітно збільшується у розмірах. Далі відбувається кілька поділів ядра, причому перший є редукційним. З дочірніх протопластів утворюється 32-64 гаплоїдні чотириджгутикові зооспори, половина з яких є чоловічими, інші - жіночими. У слизовому міхурі зооспори виходять у зовнішнє середовище, і по закінченні руху проростають у морфологічно схожі жіночі та чоловічі гаметоспорофіти.
Довгі нерозгалужені однорядні нитки характерні для гаметоспорофітів роду Ulothrix, види якого поширені у морях, хоча також досить часто зустрічаються у прісних водоймах та навіть у ґрунтах. Кожна клітина нитки (за винятком базальної, якою водорость прикріплюється до субстрату) має пристінний хлоропласт у вигляді незамкненого кільця з одним-кількома піреноїдами та одне ядро.
Вегетативне розмноження відбувається фрагментацією нитки. При нестатевому розмноженні у будь-якій клітині утворюються чотирьохджгутикові зооспори, які надалі проростають у нову нитку.
Статевий процес ізогамний. Будь-яка клітина здатна утворити велику кількість дводжгутикових ізогамет, які виходять у зовнішнє середовище і копулюють. Зигота деякий час рухається, оскільки зберігає джгутики гамет, далі зупиняється, одягається товстою оболонкою і проростає у булавоподібний одноклітинний спорофіт (стадія Codiolum). Після редукційного поділу спорофіт утворює чотирьом чотириджгутикові зооспори, що проростають у нові нитчасті гаметоспорофіти. Подібно до Chlorocystis, нитки Ulothrix є роздільностатевими.
У морського роду Spongomorpha у життєвому циклі чергуються гаметофіт та спорофіт. Гаметофіти мають вигляд кущиків, що складаються з коротких сланких ниток, видозмінених у ризоїди, та довгих розгалужених висхідних ниток, бічні гілки яких вигинаються у вигляді гачків, особливо численних при основі слані. Через гачковидні гілки нитки у нижній частині талому дуже переплутані. Таломи спонгоморфи наростають за рахунок поділу апікльних клітин ниток.
Кожна клітина має пристінний перфорований хлоропласт з багатьма піреноїдами. Як і у попередніх родів, піреноїд облямований численними зернятками крохмалю. У центральній частині клітини розташовується одне ядро.
Розмноження відбувається статевим шляхом та зооспорами. Навесні або на початку літа клітини гаметофіту продукують велику кількість дводжгутикових ізогамет. Після їх копуляції утворюється планзигота, яка після короткого періоду руху осідає на таломи макроскопічних водоростей, переважно - червоних, частково в них занурюється та одягається оболонкою. Через деякий час вона починає проростати в одноклітинний диплоїдний спорофіт (стадія Codiolum). При цьому ядро кілька разів мітотично ділиться, клітина повільно росте і набуває ценоцитної будови. Ця стадія розвитку у деяких видів спонгоморфи раніш була описана як самостійні види роду Chlorochytrium. У вигляді багатоядерного спорофіту спонгоморфа перебуває протягом літнього, осіннього та частини зимового сезону. У другій половині зими ядра спорофіту редукційно діляться і утворюється велика кількість чотириджгутикових зооспор, що надалі проростають у нові гетеротрихальні гаметофіти.
У таломах морських червоних водоростей, живих та мертвих клітинах ряски, у шкірі риб, в обгортках яєць саламандр, виявлено декілька паразитичних та ендофітних водоростей, описаних як види роду Chlorochytrium. Проте, на відміну від стадії "Chlorochytrium" у спонгоморфи, вони не утворюють багатоклітинні гаметофіти і розмножуються лише за допомогою зооспор. Деякі з цих видів, наприклад, Chlorochytrium lemnae, що розмножується за допомогою дводжгутикових зооспор х-типу, належать до Chlorophyceae. Проте не виключено, що частина хлорохітріум-подібних видів, особливо тих, що мешкають у таломах червоних водоростей, можуть виявитися ценоцитними представниками Codiolales, що втратили здатність до статевого розмноження.
У Monostroma талом гаметофіту має вигляд одношарової платівки з гофрованими краями, що має довжину до 10-20 см. Кожна клітина має пристінний хлоропласт з піреноїдом, облямованим кількома крохмальними зернятками та одне ядро. Розмноження відбувається фрагментацією таломів, акінетами, або статевим шляхом. Гамети також здатні проростати партеногенетично, як у нові гаметофіти, так і у гаплоїдні спорофіти. Таломи гаметофіту розвиваються з зооспор або партеногенетично пророслих гамет: рухлива клітина (чотириджгутикова зооспора або дводжгутикова гамета) прикріплюється до субстрату, ділиться, утворюючи спочатку двошарове скупчення клітин. Далі між першим та другим шарами заявляється порожнина, верхній шар продовжує розростатися, утворюючи велику міхуроподібну сферу, і далі відривається. Відірвана частина власне і складає одношаровий пластинчастий талом нового гаметофіту (рис. 24.42).
При статевому розмноження будь-яка клітина гаметофіту утворює велику кількість дводжгутикових гамет. Таломи у Monostroma роздільностатеві: одні гаметофіти продукують дрібні чоловічі гамети, інші - жіночі гамети, які є малорухливими, проте за розмірами помітно більші, ніж чоловічі. Статевий процес гетерогамний. Після копуляції зигота осідає на черепашки молюсків або інші кальцій-вмісні субстрати, і починає проростати у стадію Codiolum. При цьому бічний випин "свердлить" субстрат, далі розширюється у булавоподібне потовщення, у яке переходить протопласт зиготи. Спорофіт утворює численні бічні випини, і переходить у стадію, у який нагадує морські свердлячі водорості роду Gomontia. Через деякий час відбувається мейоз, і з клітини спорофіту виходить кілька чотириджгутикових зооспор, що проростають у нові гаметофіти.
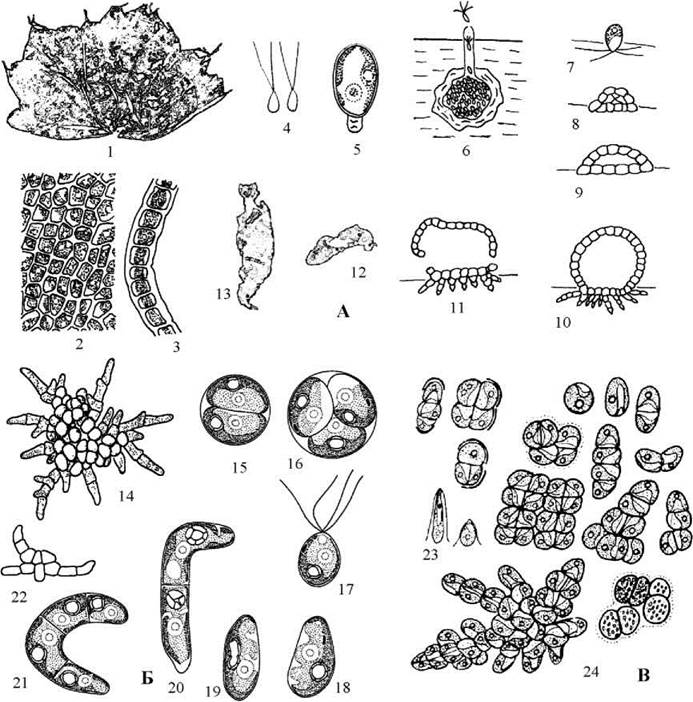
Рис. 24.42. Деякі представники Codiolales: А - Monostroma grevillei (1 - дорослий талом гаметофіту; 2, 3 - вигляд клітин з поверхні (2) та на поперечному зрізі талому (3); 4 - гамети; 5 - молодий спорофіт, стадія Codiolum; 6 - утворення зооспор дорослим спорофітом, що розвивається у черепашках молюсків; 7 - зооспора спорофіту; 8-11 - послідовні стадії розвитку талому гаметофіту у прикріпленому стані; 12, 13 - молоді таломи у вільноплаваючому стані); Б - Pseudendoclonium basiliense (14 - дорослий талом; 15, 16 - утворення зооспор; 17 - зооспора; 18-22 - послідовні стадії проростання зооспори у новий талом); В - Gloeotilopsis sarcinoidea (23 - зооспори, 24 -різноманітність форм талому (1-3, 12, 13 - за Виноградова, 1979; 4, 6 - за Kornmann, 1962; 14, 22 - за Vischer, 1926; 15-21 - за Ettl, Gartner, 1995; 23, 24 - за Lukesova, 1991).
Якщо гамети проростають партеногенетично, вони все одно дають початок стадії Codiolum. Проте в цьому випадку спорофіт є гаплоїдним, на відміну від "Codiolum", що утворюється після статевого процесу. Існує припущення, що морські свердлячі водорості роду Gomontia можуть походити від видів подібних до Monostroma, які втратили стадію гаметофіту.
Молекулярні дослідження показують, що в межах порядку існує тенденція переходу до прісноводного та наземного способів життя, яка пов"язана з вторинною втратою здатності до статевого розмноження. Перехід до виключно нестатевого розмноження відбувався як через втрату здатності до утворення гамет, так і через втрату гаметами здатності до копуляції та переходу до партеногенезу. Наприклад, за результатами аналізу ядерного гену, що кодує 18S РНК, у Codiolalaes входять агамні ґрунтові та прісноводні водорості родів Pseudoendoclonium та Gloeotilopsis, що демонструють відповідно, перший та другий варіанти агамності.
Pseudoendoclonium має таломи у вигляді коротких розгалужених ниток без чіткої диференціації на сланку та висхідну частину. Кожна клітина містить одне ядро, пристінний перфорований щілинами хлоропласт з піреноїдом. Розмноження відбувається лише фрагментацією таломів та чотириджгутиковими зооспорами. Gloeotilopsis за будовою слані та клітин подібний до попереднього роду, але розмножується за допомогою дводжгутикових зооспор. За всіма відомими сьогодні варіантами молекулярно- філогенетичних реконструкцій види останнього роду походять від форм зі статевим процесом, чотирьохджгутиковими зооспорами та гаметами, здатними до партеногенетичного розвитку у нові гаметоспорофіти.
Acrosiphonales. Порядок є філогенетично близьким до Codiolales, від якого відрізняється сифонокладальним типом структури та відсутністю субмікроскопічних лусочок на поверхні монадних клітин. Як і кодіолальні водорості, акросифональні мають гаплодиплофазний життєвий цикл з гетероморфною зміною поколінь.
Таломи гаметоспорофітів мають вигляд простих або розгалужених однорядних ниток. Кожна клітина містить сітчастий хлоропласт з багатьма піреноїдами, облямованими численними крохмальними гранулами та кілька ядер. Наростання талому відбувається внаслідок особливого варіанту сегрегативного поділу клітин: перед початком цитокінезу ядра мігрують у площину, де буде утворюватися клітинна перегородка, далі один раз синхронно діляться, а після завершення формування поперечної стінки безладно розташовуються у протопластах дочірніх клітин. Спорофіти одноклітинні, і подібно до попереднього порядку, являють собою стадію Codiolum.
У морської водорості Urospora гаметоспорофіти нерозгалужені і мають вигляд довгих ниток шириною до 0.1 мм. Нитки прикріплюються до субстрату за допомогою базальних клітин, видозмінених у ризоїди. Кожна клітина нитки, за вийнятком базальної, здатна утворювати або дводжгутикові анізогамети, або чотириджгутикові зооспори (рис. 24.43).
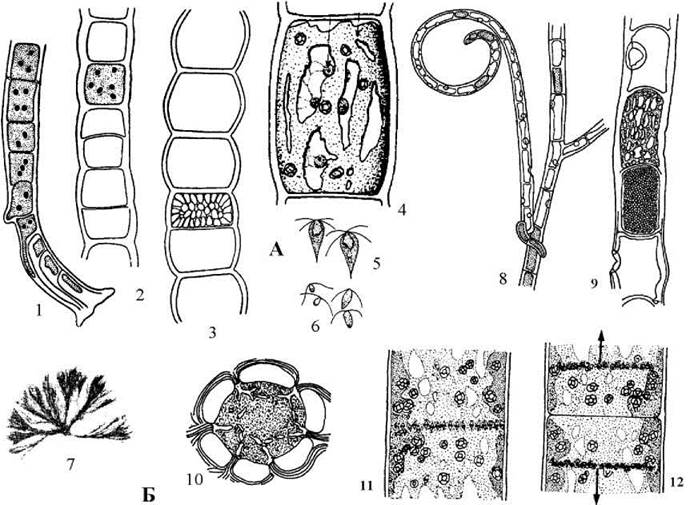
Рис. 24.43. Деякі представники Acrosiphonales: А - Urospora: 1-3 - гаметоспорофіт: базальна (1), центральна вегетативна (2) та репродуктивна (3) частини гаметоспорофіту; 4 - вегетативна клітина з багатьма ядрами, сітчастим хлоропластом з піреноїдами; 5 - зооспори; 6 - чоловічі та жіночі гамети; Б - Acrosiphonia: 7 - зовнішній вигляд талому; 8 - фрагмент талому з порожніми гаметангіями; 9 - порожні гаметангії з отвором для звільнення гамет, гаметангій та вегетативна клітина; 10 - ультратонка будова піреноїду; 11, 12 - мітоз та цитокінез: ядра розташовуються у площині майбутньої клітинної перегрородки (11), по завершенні мітозу та цитокінезу синхронно мігрують вглиб клітини (12) (1-3, 7-9 - за Виноградова, 1979; 4-6, 10 - за Lokhorst, Trask, 1981; 11, 12 - за Kornmann, 1965).
Життєвий цикл Urospora такий самий, як і у Ulothrix. Гамети уроспори здатні також партеногенетично проростати в гаплоїдний гаметоспорофіт.
Кущисті таломи з розгалужених ниток характерні для сифонокладальних водоростей з роду Acrosiphonia. Будова клітин, цитокінез, розмноження, життєвий цикл, місцезростання у видів цього роду практично ідентичні до Urospora.
Ulvales. До порядку входить біля 200 видів макроскопічних водоростей з гетеротрихальною або примітивною тканинною будовою, що мешкають переважно у морях. Філогенетично порядок є близьким до Codiolales та Acrosiphonales, від яких відрізняється відсутністю у життєвому циклі стадії Codiolum, та наявності ізоморфної зміни поколінь. Подібно до Acrosiphonales, монадні клітини ульвальних водоростей позбавлені субмікроскопічних лусочок.
Прикладом гетеротрихальної ульвальної водорості є Acrochaete. Види цього роду є морськими перифітонними організмами. Таломи гаметофіту та спорофіту морфологічно не відрізняються між собою. Вони сланкі, складаються з розгалужених одноядерних ниток. У центральній частині нитки частково зростаються, утворюючи неправильну псевдопаренхіматозну платівку. Від платівки відходять короткі ризоїдальні нитки, за допомогою яких водорість прикріплюється до субстрату, та асиміляторні нитки, що частково піднімаються над субстратом. Інколи асиміляторні нитки закінчуються одноклітиннами безбарвними волосками (хетами), бульбоподібно розширеними при основі. Кожна клітина (крім хет) має одне ядро, пояскоподібний хлоропласт з одним або кількома піреноїдами, облямованими двома шкарлупками крохмалю. У клітин асиміляторних ниток хлоропласт великий, виповнює більшу частину протопласту, у ризоїдальних клітинах хлоропласт здебільшого редукований до невеликої платівки (рис. 24.44).
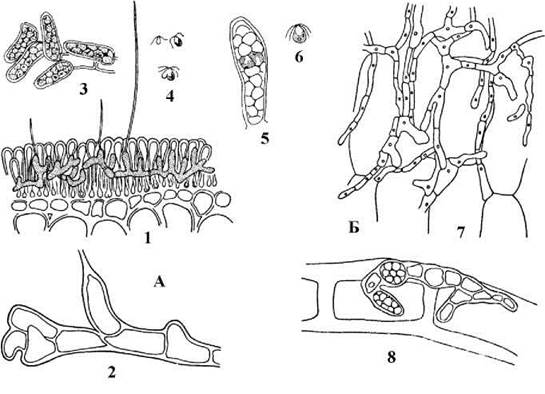
Рис. 24.44. Гетеротрихальні представники Ulvales: А - Acrochaete: 1 - талом у тканині бурої водорості Chorda; 2 - фрагмент нитки з волоском; 3 - гаметангії; 4 - гетерогамети та їх копуляція нитки; 5 - зооспорангій; 6 - зооспора; Б - Entocladia: 7 - талом на поверхні морської водорості-макрофіту; 8 - фрагмент талому з зооспорангіями (за Виноградова, 1979).
Розмноження відбувається фрагментацією ниток, зооспорами або статевим шляхом. У клітинах асиміляторних ниток спорофіту після редукційного поділу утворюються чотириджгутикові зооспори, що через отвір в оболонці виходять у зовнішнє середовище і проростають у гаметофіти. Гаметофіти однодомні - різні клітини одного й того ж талому перетворюються на чоловічі або жіночі гаметангії. Після виходу гамет у зовнішнє середовище відбувається гетерогамний статевий процес і зигота без періоду спокою проростає у новий спорофіт.
Види роду Entocladia за морфологією та життєвим циклом нагадують Acrochaete, проте не мають хет і зустрічаються не тільки у морях, але й у прісних водоймах, звичайно як епіфіти інших багатоклітинних зелених водоростей (зокрема, Cladophora, Rhizoclonium, Oedogonium).
У роду Ulva - зеленого морського салату - таломи паренхіматозні, мають вигляд двошарової платівки з гофрованими краями, довжина якої складає кілька сантиметрів. Кожна клітина містить одне ядро, пристінний пластинчастий хлоропласт з одним- кількома піреноїдами. Таломи гаметофітів розвиваються з чотириджгутикових зооспор, спорофітів - з щойно утворених зигот, що зберігають джгутики гамет (планзигот): клітина прикріплюється до субстрату, втрачає джгутики і ділиться спочатку у поперечній площині, утворюючи однорядну нерозгалужену первинну нитку. Далі клітини нитки діляться повздовжньо-радіально, утворюючи одношарову трубку з порожниною в центрі. Стінки трубки змикаються, і крайові клітини продовжують ділитися, внаслідок чого утворюється двошарова платівка зі складчастими краями (рис. 24.45).
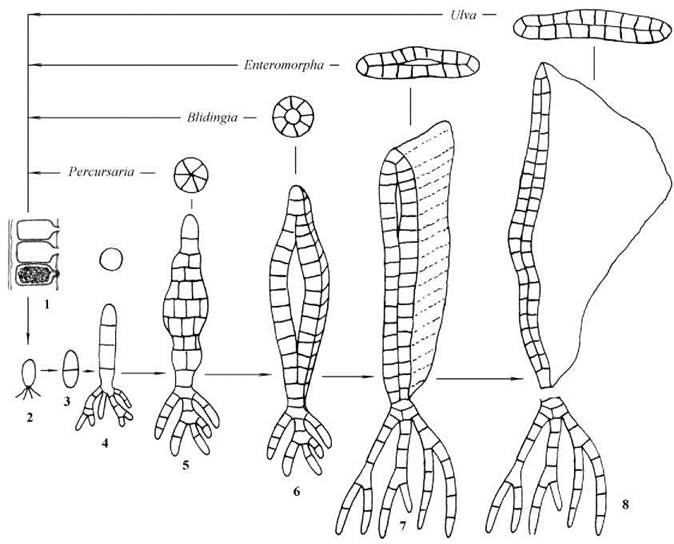
Рис. 24.45. Розвиток талому різних ульвальних водоростей з паренхіматозним типом морфологічної структури тіла: 1 - зооспорангії або гаметангії; 2 - зооспора або планзигота; 3 - початкова стадія розвитку талому; 4 - стадія однорядної нитки; 5 - стадія багаторядної нитки; 6 - трубчаста стадія з порожниною; 7 - часткове зростання стінок трубки з утворенням платівки; 8 - двошарова платівка, що відривається від сланкої частини талому.
Життєвий цикл гаплодиплофазний, з правильною ізоморфною зміною поколінь та споричною редукцією кількості хромосом.
У близьких до Ulva родів Enteromorpha, Percusaria, Blidingia, Ulvaria таломи видовжено-пластинчасті, трубчасті, або мають вигляд багаторядних ниток чи одношарових платівок (рис. 24.46). За типом життєвого циклу, будовою клітин та розмноженням всі ці роди подібні до Ulva. Відміни полягають у тому, на якій стадії припиняється розвиток талому (рис. 24.45).
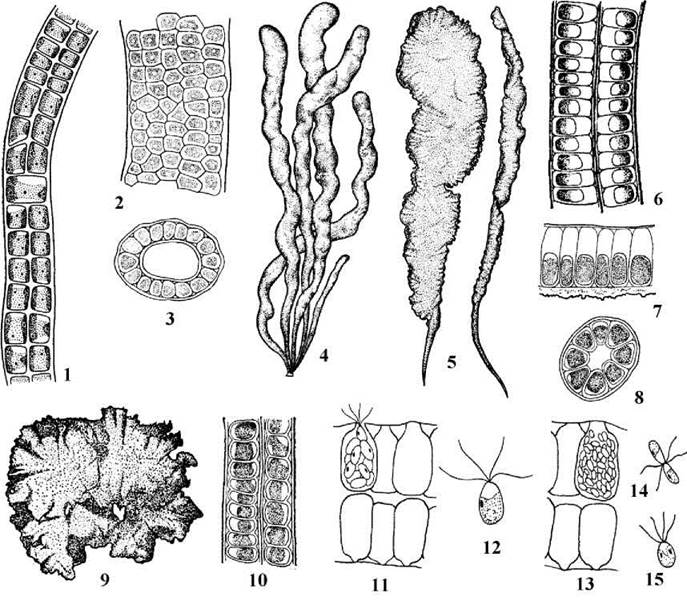
Рис. 24.46. Представники Ulvales з паренхіматозним типом морфологічної структури. 1 - Percursaria; 2, 3 - Blidingia: поверхня (2) та поперечний зріз (3) талому; 4-8 - Enteromorpha: трубчасті (4) та пластинчасті (5) таломи, поперечні зрізи талому в зоні двошарової платівки (6), середній частині з великою порожниною (7), базальній частині (8); 9-15 - Ulva: зовнішній вигляд (9) та поперечний зріз (10) талому, зооспорангій (11), зооспора (12), гаметангій (13), копуляція ізогамет (14), планзигота (15) (1, 6 - за Виноградова, 1979; 3-5, 7 - 10 - за Мошкова, 1979; 11-15 - за Koeman, van den Hoek, 1981).
Так, у Enteromorpha розвиток талому припиняється тоді, коли стінки трубки змикаються у верхівковій частині. Внаслідок цього молода водорість має вигляд кишки (звідси її народна назва - кишечниця), а доросла у нижній частині має вигляд одношарової трубки з порожниною, у верхній - двошарової видовженої платівки.
У Blidingia розвиток завершується на стадії трубки, причому її стінки не змикаються. Таломи Percusaria стають зрілими одразу після поздовжньо-радіального поділу клітин первинної нитки, порожнина звичайно не утворюється; через це водорість набуває вигляду багаторядної нитки. Рід Ulvaria проходить стадію первинної нитки та трубки. Далі верхня частина трубки розривається, розпрямляється, клітини продовжують ділитися, внаслідок чого при основі талому зберігається трубка, проте вище утворюється одношарова платівка; якщо така платівка відривається від нижньої частини, вона набуває такого вигляду, як і доросла Monostroma.
Представники цих родів є типовими мешканцями морських узбережь, проте окремі види Enteromorpha та Percusaria (P.percusa, E.intestinalis, E.prolifera) зустрічаються також у прісних континентальних водоймах.
Cladophorales (Siphonocladales). Порядок об’єднує більш 400 видів з 32 родів. Переважна більшість кладофоральних водоростей за зовнішнім виглядом нагадує представників Acrosiphonales, проте філогенетично являє собою чітко відокремлену групу. Від інших порядків ульвофіцієвих Cladophorales відрізняється за цілим комплексом як біохімічних, так і цитологічних ознак.
У кладофоральних водоростей виявлено додатковий специфічний ксантофіл сифоноксантин, що зустрічається також у празинофіцієвих та сифонофіцієвих водоростей. Клітинні оболонки побудовані з паралельно розміщених фібрил целюлози, що синтезується лінійними термінальними ферментними комплексами (рис. 24.47). Такі комплекси, крім Cladophorales, виявлені тільки у деяких видів роду Chlorella. У складі оболонок відсутні пектинові речовини. Через це поверхня клітин звичайно шорстка, шарувата, і є зручним субстратом для прикріплення різноманітних епіфітних водоростей та найпростіших.
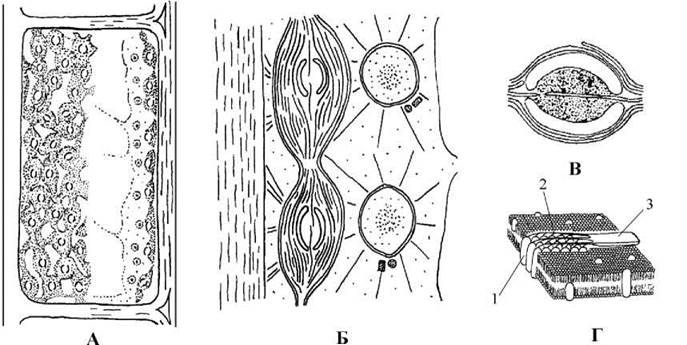
Рис. 24.47. Кладофоральні водорості: А - клітина з поверхні (ліворуч) та в оптичному розрізі (праворуч); Б - схема будови протопласту; В - білентикулярний піреноїд; Г - лінійний термінальний ферментний комплекс (1 - плазмалема, 2 - лінійна група ферментативних субодиниць, 3 - мікрофібрила целюлози) (А, Б - ориг.; В - за Hori, Ueda, 1975; Г - за Itoh, Brown, 1984).
У монадних клітин верхівкова перетинка, що з’єднує базальні тіла, є не гладкою, а поперечно-смугастою, подібною до требуксієфіцієвих водоростей. Вегетативні клітини мають багато ядер, пристінний сітчастий хлоропласт або велику кількість дрібних дископодібних хлоропластів, з'єднаних між собою дуже тонкими плазмадесма-подібними тяжами; у хлоропластах розташовуються дрібні піреноїди. Кожний піреноїд в екваторіальній площині пронизаний тилакоїдами і поділений на дві напівсфери, навколо яких відкладається суцільна напівсферична платівка крохмалю (рис. 24.47).
Протопласт сифонокладальних водоростей мають досить складну архітектуру: хлоропласти "заякорені" біля певних ділянок плазмалеми групами мікротрубочок; під кожною відособленою ділянкою хлоропласту (або під окремим хлоропластом) розташовується одне ядро. Воно, в свою чергу, оточено радіально розташованими мікротрубочками, частина яких фіксується на зовнішній мембрані хлоропласту.
Мітози у різних ядрах однієї клітини відбуваються синхронно, проте цитокінез та каріокінез між собою не зкоординовані і ядра ніколи не вистроюються у площині утворення поперечної перегородки. При мітозі спостерігаються не окремі хромосоми, а одна щільна хроматинова платівка.
При формуванні зооспор або гамет протопласт материнської клітини одночасно розпадається на велику кількість одноядерних дочірніх протопластів майбутніх спор або гамет. При цьому плазмалеми дочірніх клітин утворюються з пухирців комплексу Гольджі без помітної участі плазмалеми материнської клітини.
Життєвий цикл гаплодиплофазний, з ізоморфною зміною поколінь. Як і у попередніх порядках, зооспори переважно чотириджгутикові, гамети - дводжгутикові.
Найпростіші представники порядку мають таломи у вигляді системи розгалужених міхурів (рід Valonia), у більш складних форм таломи складаються з центральної вісі та системи з'єднаних з нею бічних сегментів (Siphonocladus). У деяких представників бічні сегменти зростаються між собою, утворюючи псевдопаренхіматозні структури (Anadyomene) (рис. 24.48). Всі ці представники зустрічаються у морях тропічних та субтропічних широт.
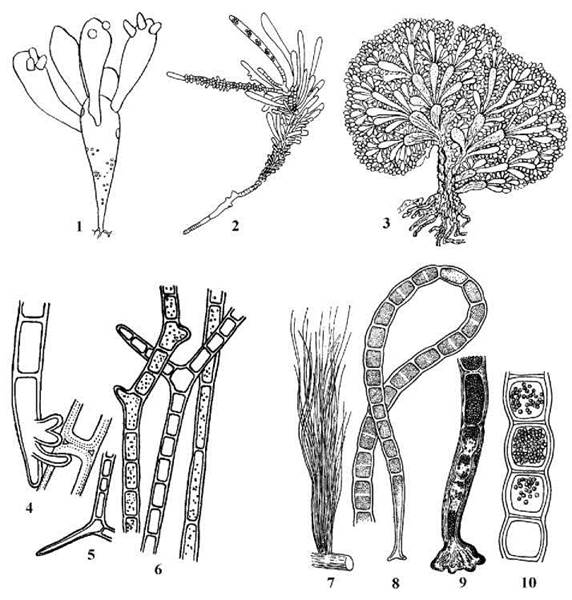
Рис. 24.48. Деякі представники Cladophorales. 1 - Valonia; 2 - Siphonocladus; 3 - Anadyomene; 4-6 - Rhizoclonium: базальна частина талому (4), коротка бічна гілка (5), фрагменти ниток (6); 7-10 - Chaetomorpha: зовнішній вигляд (7), базальні та центральна частини нитки (8, 9), фрагмент нитки зі спорангіями (10) (1 - за Виноградова, 1977; 2 - за Vickers, Shaw, 1908; за Масюк, 1993; 4-10 - за Мошкова, 1979).
Значна кількість кладофоральних має таломами у вигляді нерозгалужених ниток або розгалужених кущиків. Представники цієї групи поширені як у морях, так і у прісних континентальних водоймах.
Рід Rhizoclonium має вигляд довгих нерозгалужених ниток. Кожна клітина містить незначну кількість ядер - звичайно, не більш чотирьох. Молоді особини ведуть прикріплений спосіб життя, утримуючись на субстраті за допомогою безбарвних ризоїдів. Дорослі нитки звичайно вільноплаваючі, їх скупчення утворюють жорсткі неслизькі тини, в яких ширина клітин складає 30-50 мкм, а довжина окремої нитки - до кількох метрів. Спорофіт ризоклоніуму продукує дводжгутикові зооспори, гаметофіт - дводжгутикові анізогамети. У прісних водоймах найпоширенішим видом є R.hieroglyphicum, у морях - R.riparium.
Таломи Chaetomorpha також нерозгалужені і ниткоподібні. Проте кількість ядер у клітині складає кілька десятків, ширина ниток - 0.1-0.3 мм. Нестатеве розмноження відбувається чотириджгутиковими зооспорами та акінетами, статевий процес - за допомогою дводжгутикових ізогамет. Найпоширеніший представник роду - Ch.linum - мешкає у морях та приморських лиманах.
Провідним родом порядку є Cladophora. Таломи кладофори кущикоподібні, складаються з багатоядерних клітин, що утворюють системи розгалужених ризоїдальних та висхідних ниток. Клітини звичайно вкриті товстими шаруватими оболонками, до яких прикріплюються численні епіфітні організми.
Розмноження відбувається фрагментацією таломів, зооспорами, акінетами та за допомогою ізогамного статевого процесу.
У верхівкових клітинах спорофіту відбувається мейоз, після якого утворюється велика кількість чотириджгутикових зооспор. Через отвір в оболонці спорангію зооспори виходять у зовнішнє середовище і проростають у гаметофіт. Гаметофіти кладофори є роздільностатевими. Чоловічі та жіночі гамети морфологічно не відрізняються між собою, проте різняться фізіологічно та за ультратонкою будовою апікальної частини клітини. Так, жіночі гамети на верхівці під плазмалемою мають спеціальний ковпачок, який при копуляції приймає участь в утворенні кон’югаційного каналу. Хоча будь-яка вегетативна клітина висхідної частини талому здатна перетворитися у гаметангій, проте процес утворення гамет, як правило, починається у верхівкових клітинах талому. Через отвір в оболонці гамети звільняються і копулюють з гаметами особин іншого статевого знаку. Після копуляції зигота осідає на субстрат і без періоду спокою починає проростати у новий спорофіт.
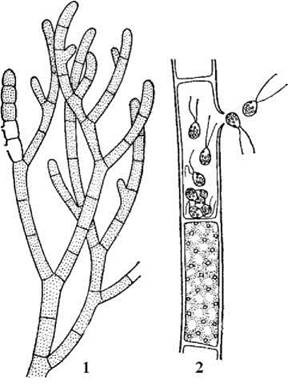
Рис. 24.49. Cladophora: фрагмент талому (1), зооспорангій та вихід зооспор (2) (за Виноградова, 1977).
Гамети кладофори (а також і деяких інших кладофоральних) здатні проростати партеногенетично, даючи початок як спорофітам, та і гаметофітам. Ця особливість зумовлює існування в межах роду досить різноманітних варіантів конкретних життєвих циклів, які, проте, є похідними від основного, гаплодиплофазного циклу зі споричною редукцією.
Найбільш поширеним видом морів помірних та теплих широт є Cladophora vagabunda. У прісних водоймах видом з найбільшим траплянням є C.glomerata. Морфологічно ці два види не відрізняються між собою, але перший має гаплодиплофазний життєвий цикл зі зміною поколінь, тоді як у другого статевий процес відсутній, а нестатеве розмноження відбувається дводжгутикомими зооспорами. Спеціальні дослідження показали, що зооспори C.glomerata вірогідно, є гаметами, що втратили здатність до копуляції, проте зберегли здатність до партеногенетичного розвитку. Таким чином, вегетативні таломи C.glomerata - це гаплоїдні гаметофіти.
З Cladophora та Rhizoclonium в деяких країнах виготовляють папір, а на Близькому Сході місцеві жителі вживають кладофору в їжу.
Trentepohliales. Порядок включає виключно наземні гетеротрихальні водорості, що, подібно до кладофоральних, мають цілий комплекс специфічних ознак.
Яскравою біохімічною особливістю є здатність накопичувати у вегетеативних клітинах велику кількість гематохрому, завдяки чому таломи трентеполіальних звичайно забарвлені у жовто-гарячий колір. Цей гематохром являє собою β-каротин, розчинений в олії та велика кількість полігідроксиспиртів - поліолу та альдітолу. Останні виявлені тільки у представників цього порядку. Інша особливість - це наявність в оболонці шару спорополеніну. Вважають, що гематохром підвищує в’язкість цитоплазми та захищає клітини від дії інтенсивного світла та надмірної втрати вологи, а спорополелін надає оболонкам хімічної стійкості та запобігає механічним ушкодженням.
Цитологічні особливості пов’язані з фотосинтетичним апаратом, цитокінезом та цитоскелетом. Так, у трентеполіальних відсутні піреноїди, крохмаль синтезується у незначній кількості і відкладається у вигляді дрібних гранул у стромі хлоропласту.
Цитокінез відбувається за участю фрагмопласту: у телофазі між мікротрубочками веретена у площині майбутньої клітинної перегородки накопичуються пухирці комплексу Гольджі, що містять мікрофібрили целлюлози. Ці пухирці зливаються як між собою, так і з плазмалемою зачаткової кільцевої борозни, обумовлюючи і ріст кільцевої борозни, і утворення клітинної платівки зі справжніми порами та плазмадесмами. Цей тип цитокінезу подібний до цитокінезу харофіцієвих та кон’югат, проте мітоз, на відміну від двох останніх класів, закритий.
Трентеполіальні мають специфічний джгутиковий апарат: на кожному джгутику є однобічний кіль, кожний джгутиковий корінь має подвійну кількість мікротрубочок, і тому загальна формула кореневої системи описуються на формулою 3-2-3-2, як у інших порядків класу, а 6-4-6-4. На верхівках тонких мікротубулярних коренів наявна колончаста структура, що нагадує багатошарову структуру деяких празинофіцієвих та харофіцієвих. Верхня зв’язка базальних тіл гладка, проте, на відміну від інших порядків, розвинена погано і розглядається як редукована.
Таломи трентеполіальних водоростей мають вигляд розгалужених ниток або одно- чи багатошарових паренхіматозних платівок.
Розмноження відбувається фрагментацією ниток, акінетами, чотириджгутиковими зооспорами та статевим шляхом. Життєвий цикл гаплодиплофазний, з ізоморфною зміною поколінь, причому зооспори утворює як гаметоспорофіт, так і спорофіт. Статевий процес ізогамний, здійснюється дводжгутиковими гаметами, які, звичайно, здатні також проростати партеногенетично.
Всі трентеполіальні водорості мешкають в аєрофітоні і є або епіфітами, що розвиваються на корі дерев, або паразитами різноманітних деревних пород.
Найбільш поширеним є рід Trentepohlia, види якої часто зустрічаються на корі листяних порід дерев, викликаючи червоне аерофітне "цвітіння". Таломи Trentepohlia мають вигляд коротких розгалужених сланких ниток, висхідні нитки редуковані (рис. 24.50). У багатьох видів кінцеві клітини помітно збільшені за розміром і являють собою або зооспорангії, або гаметангії. Звичайно після приготування водних препаратів можна спостерігати, як протягом кількох хвилин з кінцевих клітин виходять гамети або зооспори.
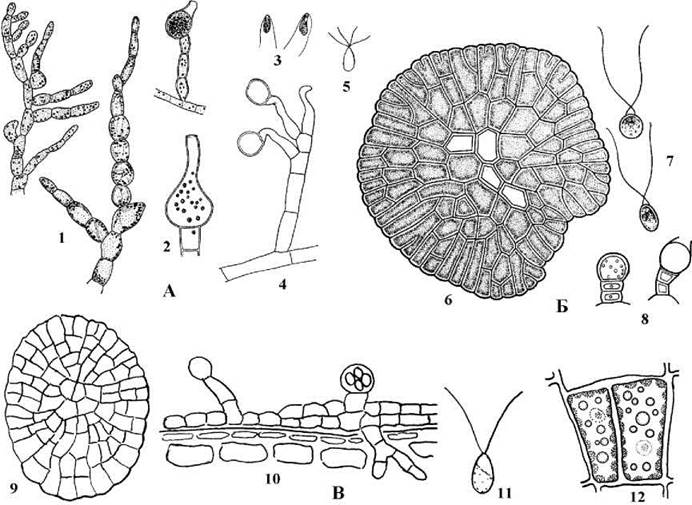
Рис. 24.50. Представники Trentepohliales. А - Trentepohlia: 1 - таломи, 2 - гаметангії, 3 - гамети, 4 - зооспорангій, 5 - зооспора; Б - Phycopeltis: 6 - талом, 7 - гамети, 8 - зооспорангії; В - Cephaleuros: 9 - талом, 10 - одно- та двошарові ділянки талому на поверхні ураженого листка, зооспорангії та ризоїдальні нитки; 11 - зооспора; 12 - вегетативні клітини (1-8 - за Мошкова, 1979; 9-12 - ориг.).
У роду Phycopeltis, види якого інколи можна знайти в оранжереях з тропічними рослинами, талом має вигляд одношарової паренхіматозної платівки, від якої відходять короткі висхідні нитки зі спорангіями на верхівці.
Епіфітна та ендофітна водорость Cephaleuros має талом у вигляді багатошарової платівки, від якої відходять не тільки висхідні нитки зі спорангіями, але й ризоїди. Цефалеврос росте на листях тропічних та субтропічних дерев та чагарників; відомі випадки, коли види цього роду спричинювали епіфітотії на плантаціях кофе, цитрусових, чаю.
За наявністю фрагмопласту та колончастої структури на джгутикових коренях трентеполіальні нагадують примітивних представників Charophyceae, проте молекулярно- філогенетичні реконструкції показують, що цей порядок все ж таки являє собою самостійну гілку Ulvophyceae, хоча й досить спеціалізовану.