Альгологія - Костіков І.Ю. - 2009-2013
Розділ 24. Зелені водорості – Chlorophyta
24.2. Хлорофітна лінія
24.2.4. Клас Siphonophyceae
Об'єднує морські водорості виключно з сифональним типом морфологічної структури тіла. Подібно до Ulvophyceae, клас представляє лінію еволюції зелених водоростей, у яких коренева система джгутиків хрестоподібна, базальні тіла зміщені проти годинникової стрілки, а фікопласт відсутній. Найдавніші викопні рештки сифонофіцієвих відомі з докембрію і датуються віком 1.2 млрд років.
Клас нараховує біля 200 сучасних та більш 300 викопних видів.
Ознаки та характеристика класу
Біохімічні особливості
Характерною біохімічною ознакою класу є наявність специфічних ксантофілів сифонеіну та сифоноксантину. Як запасна поживна речовина накопичується не тільки крохмаль, але й полімер фруктози полісахарид інулін. Целюлоза клітинних покривів складається з маннану або ксилану, а не з глюкану, як це має місце в інших класах Chlorophyta (у бриопсидальних водоростей у слідових кількостях виявлені також і глюкани, проте мікрофібрил целюлози у клітинних стінках вони не утворюють).
Цитологічні особливості
Таломи мають сифональну організацію, за винятком молодих стадій. У центрі талому знаходиться велика вакуоля з клітинним соком, на периферії - тонкий шар цитоплазми з хлоропластами, ядрами та іншими органелами (рис. 24.51).
Специфічна особливість сифонофіцієвих - наявність впорядкованого руху цитоплазми. Вона рухається разом з хлоропластами та ядрами, за певними напрямками. Процес можна спостерігати у звичайний оптичний мікроскоп. Швидкість руху складає 60120 мкм/хв. Рух відбувається за участю актину та тубуліну. Додавання у середовище специфічного інгібітору актину - цитохалазину В - призводить до припинення руху. Участь мікротрубочок у цьому процесі доведена методами імунофлюоресцентної мікроскопії. Активний рух цитоплазми виявлений також у харофіцієвих водоростей та у вищих рослин.
Клітини сифонофіцієвих великі (від кількох мм до кількох см), і досить часто пошкоджуються тваринами або механічно. Проте пошкодження не призводить до загибелі особини, оскільки існує механізм швидкого заживлення ран за допомогою спеціальних пробок. В місці пошкодження протягом кількох секунд утворюється сітка з актину, далі з вакуолі вичавлюється попередник пробки, що накладається на актинову сітку та полімеризується. Після цього до ушкодженої ділянки транспортується целюлозний матеріал, який відкладається у зоні пошкодження і завершує формування пробки.
Клітинні покриви у багатьох представників накопичують вапно, що відкладається на поверхні клітин у формі арагоніту. Субмікроскопічні лусочки на вегететивних та монадних клітинах відсутні.
Ядерний апарат та особливості мітозу. В онтогенезі сифонофіцієві проходять стадії з двома морфологічно відмінними типами ядер - первинними та вторинними. Первинне ядро велике (100-300 мкм у діаметрі), звичайно займає у клітині фіксоване положення і розвивається з ядра зиготи. Назовні від ядерної оболонки первинного ядра розташовується тонка зона проміжної цитоплазми, що позбавлена органел; за нею - система дрібних вакуолей, що оточують ядро з проміжною зоною - т.з. лакунарний лабіринт. Далі знаходиться основна цитоплазма з хлоропластами, мітохондріями, білок-синтезуючим апаратом і т.п. Зв'язок між проміжною та основною цитоплазмою здійснюється з'єднувальними цитоплазматичниим каналами. Під ядерною оболонкою первинного ядра розташовується периферичний фібрилярний ретикулюм (у дазикладальних), а над нею - щільні перинуклеарні тіла (у бриопсидальних). В ядерній оболонці дуже багато пор типової для еукаріот будови з вісьмома периферичними та однією центральною глобулами (рис. 24.51).
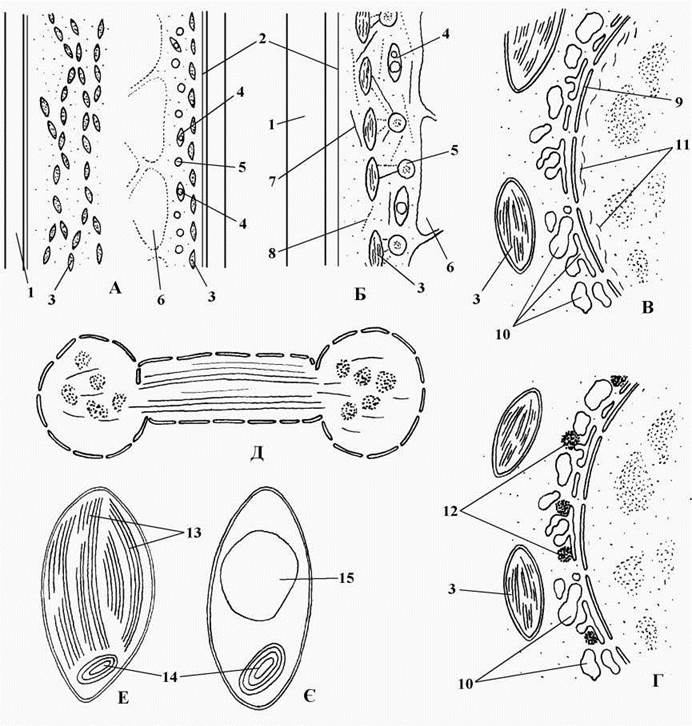
Рис. 24.51. Деякі цитологічні особливості сифонофіцієвих водоростей. А - загальний вигляд клітини (праворуч - з поверхні, ліворуч - в оптичному розрізі); Б - схема зв'язків між мікротрубочками й актиновими мікрофіламентами та основними органелами у периферичному шарі цитоплазми клітини; В, Г - схема будови первинного ядра у дазикладальних (В) та бриопсидальних (Г) водоростей; Д - вторинне ядро у телофазі; Е - хлоропласт; Є - амілопласт. 1 - клітинна оболонка, 2 - плазмалема, 3 - хлоропласт, 4 - амілопласт, 5 - вторинне ядро, 6 - вакуоля з клітинним соком, 7 - мікротрубочка, 8 - актиновий мікрофіламент, 9 - ядерна оболонка, 10 - лакунарний лабіринт у проміжній зоні первинного ядра, 11 - периферичний мікрофібрилярний ретикулюм, 12 - перинуклеарні тіла, 13 - тилакоїди, 14 - тилакоїд-організуюче тіло, 15 - крохмаль (А - ориг., Б, В, Г - схематизовано за Menzel, Schliwa, 1986; Burr, West, 1971; Hori, 1981; Hori, Ueda, 1975).
Вторинні ядра дрібні (5-15 мкм у діаметрі), вони утворюються з первинного ядра після мейозу та серії послідовних мітозів. Вторинні ядра не оточені проміжною цитоплазмою та лакунарним лабіринтом, звичайно за допомогою мікротрубочок та актинових фібріл пов'язані з хлоропластами і разом з ними циркулюють по клітині у процесі руху цитоплазми.
Мітоз у сифонофіцієвих в цілому подібний до кладофоральних водоростей: він закритий, веретено у телофазі не руйнується, а ядра на цій фазі мають характерну гантелеподібну форму (рис. 24.51). Проте на відміну від Cladophorales, центриолі звичайно відсутні. У деяких бриопсидальних водоростей при утворенні гамет мітози ацентричні, проте мітози, що відбуються у процесі наростання талому, здійснюються за участю центриолей. В останньому випадку центриолі утворюються поблизу ядра de novo, на початку мітозу реплікуються, мігрують до полюсів ядра, де приймають участь у формуванні веретена поділу. По закінченні мітозу центриолі зникають.
Фотосинтетичний апарат представлений численими дрібними дископодібними парієтальними хлоропластами, переважно без піреноїдів. У галімедальних та дихотомосифональних водоростей талом має два типи пластид - забарвлені хлоропласти та безбарвні амілопласти. Амілопласти не приймають безпосередньої участі у фотосинтезі, а виконують функції накопичення запасних полісахаридів. У багатьох синофіцієвих у хлоропластах та амілопластах є особливі тилакоїд-організуючі тіла (рис. 24.51).
Монадні стадії представлені дво- та чотириджгутиковими гаметами та стефаноконтними зооспорами (останні виявлені лише у бриопсидальних водоростей - див. нижче). Базальні тіла гамет орієнтуються за типом 11-5, глибоко перекриваються одне з одним, у латеральній позиції розгорнуті під великим кутом (біля 180о) (рис. 24.52).
Система джгутикових коренів хрестоподібна. Формула коренів в цілому відповідає формулі 4-2-4-2, проте кількість мікротрубочок у коренях варіює як у межах одного порядку, так і навіть у різних гамет однієї особини. Наприклад, кількість мікротрубочок джгутикових коренів у чоловічих гамет Pseudobryopsis описується формулою 4-2-4-2, у жіночих - 4-3-4-3. Верхня з'єднувальне волокно гладке, на відміну від Ulvophyceae воно не суцільне, а складається з двох половин, що сполучаються смугастою перетинкою. Термінальні ковпачки на базальних тілах є (Bryopsidales, Halimedales) або відсутні (Dasycladales), колончаста структура не виявлена.
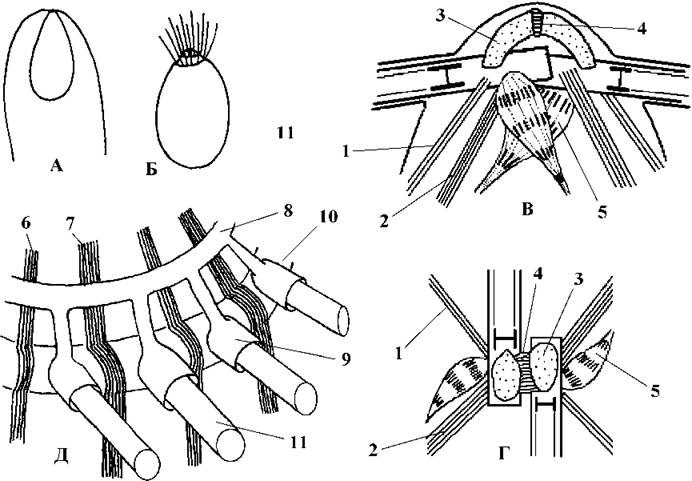
Рис. 24.52. Джгутикові стадії та джгутиковий апарат сифонофіцієвих водоростей. А - типовий план будови гамет; Б - стефаноконтна зооспора; В, Г - коренева система джгутиків гамет та нестефаноконтних зооспор: 1 - тонкий мікротубулярний корінь з двох мікротрубочок; 2 - товстий мікротубулярний корінь з чотирьох мікротрубочок; 3 - гладке верхнє з'єднувальне волокно; 4 - поперечно-смугаста перетинка з'єднувальних волокон; 5 - поперечно-смугастий додатковий мікрофібрилярний корінь; Д - коренева система джгутиків стефаноконтних зооспор: 6 - тонкий мікротубулярний корінь з чотирьох мікротрубочок; 7 - товстий мікротубулярний корінь з шести мікротрубочок; 8 - верхнє гладке з'єднувальне волокно; 9 - джгутикова муфта верхнього кільця; 10 - нижнє з'єднувальне кільце; 11 - джгутик (А, Б - за Зауер, 1977; Виноградова, 1979; В, Г - за Roberts, Stewart, Mattox, 1984; Д - за Hori, Kobara, 1982).
Типи морфологічної структури тіла
Всі сифонофіцієві водорості мають сифональний тип структури. Основним структурним елементом талому є багатоядерний сифон, що виглядає як окрема велика клітина, що не відмежована від інших сифонів клітинною перегородкою.
Таломи сифонофіцієвих в принципі відповідають двом морфологічним планам: у бриопсидальних та галімедальних тіло має вигляд тонких чи товстих ниток або шнурів, рясно або бідно розгалужених, проте не радіально-симетричних. У дазикладальних водоростей таломи складаються з центрального "стеблового" сифону та системи сифонів-мутовок, розміщених радіально на верхівці "стебла".
Сифон, що розвивається з зиготи, може тривалий час зберігати лише одне первинне ядро.
Розмноження та життєві цикли
Розмноження здійснюється за допомогою дво-, чотириджгутикових та стефаноконтних зооспор та статевим шляхом. Статевий процес ізо- або гетерогамний, і лише у одного представника (Dichotomosiphon) оогамний.
При утворенні репродуктивних клітин на спорангій або гаметангій може перетворитися або весь талом, або лише його частина. Перший шлях розвитку називають холокарпічним, другий - еукарпічним. При еукарпічному шляху розвитку сифони, де починається утворення монадних клітин, відмежоваються від основної частини талому пробками. У дазикладальних гамети утворюються лише з особливих гаметичних цист (див. нижче).
Життєві цикли гаплофазні або гаплодиплофазні, без зміни поколінь або з гетероморфною зміною. У багатьох представників відомі досить складні цикли з дигенетичним розвитком зиготи.
Система класу
Клас поділяють на чотири порядки - Bryopsidales, Halimedales, Dichotomosiphonales та Dasycladales. Перші три порядки філогенетично є досить близькими, в той час як останній являє собою специфічну, чітко відмежовану еволюційну гілку.
Сьогодні встановлено кілька фенотипічних ознак, що корелюють з поділом класу на порядки згідно до молекуляно-філогенетичних реконструкцій. Основними серед них є тип симетрії талому, наявність стефаноконтних зооспор, особливості фотосинтетичного апарату (зокрема, наявність амілопластів), спосіб утворення гамет (за холо- чи еукарпічним типом або через гаметичні цисти), тип статевого процесу, особливості організації первинного ядра, зокрема, наявність перинуклеарних тіл чи периферичного фібрилярного ретикулюму (табл. 24.6).
Таблиця 24.6. Деякі основні систематичні ознаки порядків сифонофіцієвих водоростей
Порядок |
Радіальна симетрія талому |
Стефаноконтні зооспори |
Амілопласти |
Ознака Спосіб утворення гамет |
Первинне ядро |
Статевий процес |
Bryopsidales |
- |
+ |
- |
еукарпічний |
ПФР |
гетерогамія |
Halimedales |
- |
- |
+ |
холокарпічний |
ПФР |
гетерогамія |
Dichotomo-siphonales |
- |
- |
+ |
еукарпічний |
? |
оогамія |
Dasycladales |
+ |
- |
- |
гаметичні цисти |
ПТ |
ізогамія |
Скорочення: ПФР - перинуклеарний фібрилярний ретикулюм, ПТ - перинуклеарні тіла, ? - не досліджено.
Bryopsidales. Об'єднує водорості, таломи яких не мають раділаьно-симетричної будови і являють собою одну велетеньську клітину з багатьма (інколи - до кількох тисяч) ядрами. Покриви утворені переважно ксиланами та мананами. Бриопсидальні водорості є гомопластидними організмами, їх фотосинтетичний апарат представленй лише хлоропластами; амілопласти відсутні. Особливість ядерного апарату полягає у наявності в первинному ядрі перинуклеарного фібрилярного ретикулюму, що на думку деяких дослідників, складається з великої кількості прорибосом. Центриолі звичайно відсутні, за вийнятком видів роду Pseudobryopsis, у яких про поділі дорослих ядер у пізній інтерфазі спостерігається поява центриолей, що зникають одразу по закінченні мітозу.
Характерна особливість бриопсидальних - наявність у життєвому циклі стефаноконтних зооспор (рис. 24.52). Ці зооспори мають біля 30-40 джгутиків. Базальні тіла кожного джгутика з'єднані з гладкою нижньою спіралеподібною білковою стрічкою. Вона не замкнена у кільце, робить 2-4 оберти навколо переднього кінця клітини. Крім того, кожне базальне тіло оточене фібрилярною муфтою. Муфти прикріплюються до верхньої гладкої стрічки, що походить від верхнього фібрилярного з'єднувального волокна базальних тіл. Між базальними тілами до стрічок почергово прикріплюються мікротубулярні корені, відповідно з чотирьма та шістьма мікротрубочками.
Бриопсидальні - це водорості, у яких гамети утворюються еукарпічним шляхом, тоді як зооспори - як еу-, так і холокарпічно. Розмножуються бриопсидальні фрагментацією талому, апланоспорами, стефаноконтними зооспорами та статевим шляхом.
Фрагментація відбувається шляхом відокремлення частини талому, переважно - під впливом різних механічних чинників, і в цілому спостерігається зрідка. Після відокремлення такої частини у місці розриву швидко утворюється целюлозна пробка, яка закриває місце пошкодження.
При розмноженні зооспорами та апланоспорами у одних родів (Derbesia, Bryobesia) частини сифонів відокремлюються пробкою і перетворюються на зооспорангії, що дають початок багатьом стефаноконтним зооспорами. У інших родів (Bryopsis) весь мікроскопічний спорофіт перетворюється на спорангій. Таким чином, спорофіти бриопсидальних можуть бути як холокарпічними, так і еукарпічними, тоді як гаметофіти завжди еукарпічні. В межах порядку простежується тенденція втрати здатності до нестатевого розмноження у високорозвинених форм (наприклад, кодієві водорості зооспор не мають і розмножуються лише статевим шляхом).
Статевий процес переважно гетерогамний. Життеві цикли у Bryopsidales гаплофазні, без чергування поколінь або гаплодиплофазні, з нерегулярним гетероморфним чергуванням поколінь і дигенетичним шляхом розвитку зиготи. Спорофіти проходять стадію мікроскопічної розгалуженої нитки, що отримала назву стадії "Derbesia".
Типовий представник порядку - Bryopsis - поширений у морях тропічного, субтропічного та помірного поясів (рис. 24.53). Гаметофіти цієї водорості диференційовані на сланкі ризоїдальні та висхідні асиміляторні сифони. Висхідні сифони зазвичай пірчасто розгалужені і нагадують пташине пір'я. При утворенні гамет деякі бічні сифони відокремлюються перегородкою. З сифонів, забарвлений в інтенсивно зелений колір, утворюються жіночі гаметангії, з світліших, жовтуватих сифонів - чоловічі. Далі вміст гаметангіїв розпадається на численні дводжгутикові жіночі та чоловічі гамети, що різняться за розмірами. Через отвір в оболонці гамети виходять у зовнішне середовище, де попарно копулюють, утворюючи планзиготу.
Планзигота осідає на субстрат, втрачає джгутики і без періоду спокою починає проростати у диплоїдний розгалужений ниткоподібний спорофіт, що отримав назву стадії Derbesia. Розвиток стадії Derbesia відбувається 2-4 місяці. Ядро у 20-40 разів збільшується у діаметрі, переходячи зі стадії диплоїдного зиготичного ядра у стадію первинного ядра. Далі у первинному ядрі відбувається серія мітотичних поділів, дочірні ядра зменшуються у діаметрі до 5-10 мкм. Останній поділ ядра є редукційним. Після мейозу завершується формування вторинних ядер, і спорофіт холо- або еукарпічним шляхом дає початок одноядерним стефаноконтним гаплоїдним зооспорам. Ці зооспори проростають або у нові гаметофіти, які складаються з сифонів пірчастої будови, або у нові спорофіти стадії Derbesia (рис. 24.53).

Рис. 24.53. Бриопсидальні водорості. 1-11 - стадії розвитку Bryopsis: 1 - зовнішній вигляд гаметофіту; 2 - верхівкова частина "гілки" гаметофіту; 3 - чоловічій гаметангій; 4 - чоловіча гамета; 5 - жіночій гаметангій; 6 - жіноча гамета; 7 - зовнішній вигляд спорофіту (стадія Derbesia); 8 - молодий одноядерний спорофіт з первинним ядром; 9 - спорофіт з багатьма первинними ядрами; 10 - зооспорангій з протопластами майбутніх зооспор, що мають вторинні ядра; 11 - зооспора. 12 - фрагмент талому гаметофіту Pseudobryopsis з великими вегетативними сифонами, на яких розвиваються дрібніші генеративні сифони- гаметангії (за Виноградова, 1977, 1979; Brock, Schnetter, 1997).
Цікаво, що за певних умов планзигота може безпосередньо проростати у гаметофіт, не проходячи стадію Derbesia. Проте процеси, що відбуваються з ядром при цьому шляху розвитку, залишаються не дослідженими.
Таким чином, Bryopsis має гаплодиплофазний життєвий цикл з неправильним гетероморфним чергуванням поколінь та споричною редуцією (принаймні, при типовому шляху розвитку зиготи).
Аналогічну будову та життєвий цикл має рід Pseudobrypsis, у якого, на відміну від попереднього роду, спостерігається функціональна диференціація бічних сифонів на вегетативні, що виконують лише функцію фотосинтезу, та генеративні, з яких утворюються гаметангії. Генеративні сифони мають менші розміри, і вже на ранніх фазах розвитку відокремлюються від основної частини талому поперечними перегородками.
Рід Halicystis об'єднує свердлячі водорості, що оселяються на вапнякових таломах червоних водоростей з роду Lithothamnion і частково руйнують літотамнієві вапняки. Таломи складаються з коротких сланких, ризоїдальних та міхуроподібних висхідних сифонів. З останніх утворюються гаметангії. У більшості видів цього роду життєвий цикл, подібно до попередніх родів, гаплодиплофазний, зі споричною редукцією та стадією Derbesia (рис. 24.54).
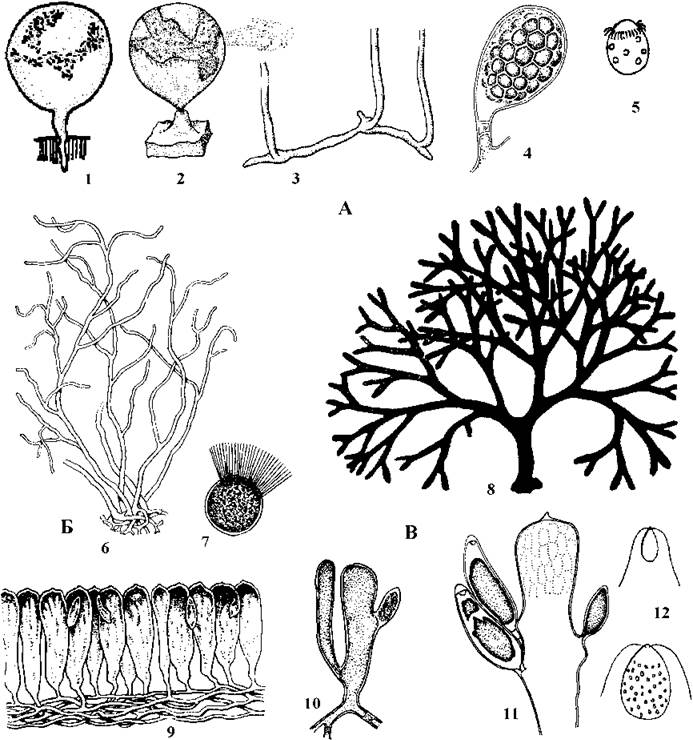
Рис. 24.54. Бриопсидальні водорості. А - Halicystis: 1 - дорослий талом гаметофіту з ризоїдом, що свердлить вапняковий субстрат; 2 - гаметангій, з якого виходять гамети; 3 - частина талому спорофіту (стадія Derbesia); 4 - зрілий зооспорангій; 5 - зооспора. Б - Derbesia: 6 - талом, 7 - зооспора. В - Codium: 8 - зовнішній вигляд талому; 9 - фрагмент талому з видовженими ризоїдальними сифонами від яких відгалужуються міхуроподібні сифони-утрикули; 10 - загальний вигляд утрикули; 11 - частина утрикули з гаметангіями; 12 - чоловічі та жіночі гамети (за Виноградова, 1977; Eckhardt, Schnetter, 1984).
У надзвичайно широко розповсюдженого в морях космополітичного виду Halicystis ovalis у життєвому циклі наявна стадія дикаріону: жіночі та чоловічі гаметофіти продукують анізогамети, що після звільнення з гаметангіїв копулюють. При копуляції цитоплазми гамет зливаються, проте каріогамія не відбувається і утворюється дикаріонтична клітина. Вона осідає на субстрат та розвивається у спорофіт, відомий під назвою "Derbesia marina". Талом цієї стадії являє собою розгалужену систему сланких та висхідних сифонів, що містять велику кількість дрібних чоловічих та жіночих ядер. Далі на довгих висхідних сифонах розвиваються короткі бічні сифони, що перетворюються на спорангії. При формуванні цих спорангіїв "чоловічі" та "жіночі" ядра зливаються, утворюючи диплоїдні зиготичні ядра. Вони одразу редукційно діляться, і протопласт клітини розпадається на велику кількість стефаноконтних гаплоїдних зооспор, що надалі проростають у нові чоловічі та жіночі гаметофіти.
У межах роду Halicystis відомі також види, у яких спорофіт відсутній, і розмноження відбувається тільки статевим шляхом, без стадії Derbesia. В цьому випадку молоді таломи диплоїдні, дорослі - гаплоїдні, що є наслідком соматичної редукції кількості хромосом. Вважають, що такі форми виникли через зміну дигенетичного шляху розвитку зиготи на моногенетичний, що призвело до втрати стадії спорофіту.
У морях теплого та помірного поясів вельми поширеними є види роду Derbesia, за яким отримали свою назву спорофіти бриопсису та галіцистісу. Талом дербезії складається з коротких сланких та висхідних ниткоподібних сифонів. Статевий процес у видів цього роду відсутній, і розмноження здійснюється лише стефаноконтними зооспорами або апланоспорами. Вірогідно, що різні види Derbesia походять від бриопсис- та галіцистіс-подібних пращурів, у яких дигенетичний цикл був порушений і здатність до самовідновлення зберіглась лише у спорофіту.
Прикладом водорості зі складно диференційованим таломом є Codium, що мешкає у теплих морях, звичайно на глибині від 5 до 40 м. В Чорному морі досить поширеним видом є Codiumvermilara. Його талом має центральну частину, що складається зі щільно переплетених видовжених ризоїдальних сифонів. На периферії від ризоїдальних сифонів відгалужуються пухирчасті вирости - утрикули, що утворюють шар, подібний до кори. Система ризоїдальних сифонів та сифонів-утрикул має вигляд товстих (до 2 см у діаметрі) дихотомічно розгалужених шнурів, поверхня яких завдяки утрикулам здається губчастою (рис. 24.54).
Codium розмножується лише статевим шляхом, статевий процес - гетерогамія. Дводжгутикові жіночі та чоловічі гамети формуються у гаметангіях, що відокремлюються від утрикул перегородками, тобто водорість є еукарпічною. Чоловічі гаметангії забарвлені у жовтуватий колір, жіночі - у темно-зелений. Після копуляції зигота без періоду спокою проростає у новий гаметофіт. Вважається, що мейоз у Codium гаметичний, і життєвий цикл, відповідно, диплофазний. Гамети здатні також проростати партеногенетично.
Halimedales. Водорості цього порядку як за фенотипічними, так і за молекулярними ознаками близько споріднені з Bryopsidales: вони не мають радіально-симетричної будови, гамети утворюються в гаметангіях, а не в гаметичних цист, фаза розвитку, на якій водорість має талом лише з первинним ядром, короткочасна, а саме первинне ядро не містить перинуклеарних тіл.
Проте Halimedales має кілька яскравих специфічних ознак. По-перше, покриви водоростей утворені ксиланом і звичайно просякнуті вапном, що відкладається на поверхні клітин у формі голок арагоніту. По-друге, галімедальні водорості гетеропластидні - крім забарвлених хлоропластів наявні також безбарвні амілопласти, що не здатні до фотосинтезу і виконують лише запасаючу функцію. По-трете, як в хлоропластах, так і в амілопластах є унікальна структура, що отримала назву тилакоїд - організуючого тіла. Вона розташовується на одному з полюсів пластиди, і являє собою систему з трьох-чотирьох замкнених у невеличкі кільця тилакоїдів, які вкладені одне в одне. По-четверте, представники порядку є холокарпічними, і при утворенні гамет талом водорості перетворюється на один велетенський гаметангій.
У галімедальних стефаноконтні зооспори відсутні. Розмноження відбувається фрагментацією таломів та статевим шляхом. Статевий процес ізогамний або гетерогамний. Галімедальні водорості мешкають переважно у тропчних морях.
Таломи галімедальних водоростей макроскопічні, складно розчленовані і нагадують невелички трав'янисті рослини. Так, Halimeda диференційована на систему сланких ризомів, ризоїди та висхідні асиміляторні "пагони", що, залежно від виду, нагадують китиці, віяла або членисті мутовки. Ризоми утворені ниткоподібним сифоном, від якого у субстрат заглиблюються рясно розгалужені сифони-ризоїди. Від ризому над кожною групою ризоїдів догори відходить група сифонів-асиміляторів, утворюючи "пагони". "Пагон" складається з багатоосьового більш-менш циліндричного "стебельця" та звичайно сплощеної або багаторазово розгалуженої верхівки. Кожна вісь "пагону" - це ниткоподібний висхідний сифон, що несе на собі утрикули (рис. 24.55). Ниткоподібні сифони щільно переплетені і з'єднуються між собою численними анастомозами. Всередині сифонів є система калозних балок, яка виконує функцію внутрішнього скелету; поверхня сифонів вкрита кристалами арагоніту, що утворюють зовнішній скелет.
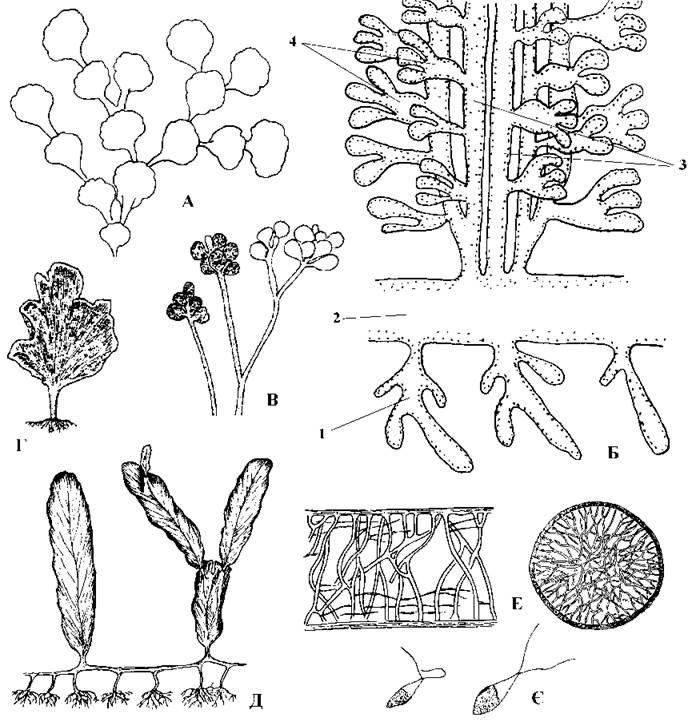
Рис. 24.55. Галімедальні водорості. А-В - Halimeda: А - зовнішній вигляд талому; Б - схема будови талому (1 - ризоїди, 2 - ризом, 3 - висхідні сифони "пагону", 4 - утрикули); В - сифони-гаметангії, що розвиваються на "пагонах", проте не відокремлюються від них перегородками; Г - зовнішній вигляд талому Udotea; Д-Є - Caulerpa: Д - зовнішній вигляд талому, Е - калозні опірні балки (поздовжній та поперечний зрізи через ризом); Є - чоловічі та жіночі гамети (за Виноградова, 1977; Meinesz, 1980; Fritsch, 1935).
Функцію фотосинтезу виконують утрикули висхідних сифонів. Вони містять численні хлоропласти та амілопласти. Вдень пластиди зосереджуються у периферичній частині утрикул, вночі вони мігрують в глибину талому і концентруються у ниткоподібних сифонах, захищених зовнішнім скелетом.
Вегетативне розмноження у галімеди відбувається при розриві ризомів. Тоді пошкоджені сифони перекриваються пробками, а ділянка талому, що відокремилась, здатна прикріпитися до субстрату і прорости у нову рослину.
При статевому розмноженні протопласт всієї рослини розпадається на велику кількість дводжгутикових жіночих або чоловічих гамет. Гамети виходять назовні і копулюють. Halimeda є роздільностатевою - чоловічі та жіночі гамети утворюються на різних рослинах. Статевий процес гетерогамний. Зигота без періоду спокою проростає в одноядерну сферичну рослину - протосферу. У протосфері ядро мітотично ділиться, утворюючи велику кількість первинних ядер. Далі протосфера утворює численні випини, розвиває систему ризомів, ризоїдів, і, нарешті, висхідних "листків". На цій стадії ядра вже вторинні. Вважають, що завершення розвитку протосфери і утворення вторинних ядер відбувається внаслідок мейозу, хоча прямі докази цього поки що відсутні.
Види Halimeda мешкають у тропічних морях і належать до рифоутворюючих водоростей. Особливо багато Halimeda у лагунах атолів - тут відмерлі таломи водорості суцільним килимом вкривають дно і утворюють особливий, спеціфічний для атолів арагонітовий пісок.
Види роду Caulerpa в цілому мають будову подібну до галімеди, але ризоми у каулерпи товсті, багатоосьові, з численними системами ризоїдів. Від ризомів відходять висхідні "пагони", що у різних видів мають різну форму. Наприклад, у Caulerpa prolifera окремий "пагон" має вигляд видовженої ланцетоподібної платівки до 30 см завдовжки. Розташування пагонів та ризоїдів на ризомі між собою не узгоджено. Вегетативне та статеве розмноження відбувається так само, як і у Halimeda.
У роду Udotea таломи мають вигляд віялоподібних платівок, закріплених у ґрунті системою сифонів-ризоїдів, причому ризомів немає. Особливістю циклу розвитку удотеї є те, що вторинні ядра утворюються після першого ж поділу первинного ядра, і, таким чином, стадія сифону з багатьма первинними ядрами відсутня.
Dichotomosiphonales. Включає лише один рід з одним видом - Dichotomosiphon tuberosum. Ця водорость, на відміну від інших представників класу, мешкає у прісних континентальних водоймах. Вона має дуже прості таломи, що утворені дихотомічно розгалуженими трубками-сифонами, закріпленими у ґрунті короткими ризоїдами, оогамний статевий процес, причому зигота проростає після періоду спокою (рис. 24.56). Проте, як і у представників попередніх порядків, у Dichotomosiphon спостерігається рух цитоплазми, базальні тіла орієнтовані за типом 11-5 і розгонуті на 180о, верхнє з'єднувальне волокно базальних тіл гладке та не суцільне (на відміну від інших сифонофіцієвих, воно складається не з двох, а з чотирьох сегментів). Кількість х-коренів дорівнює 4 або 5.
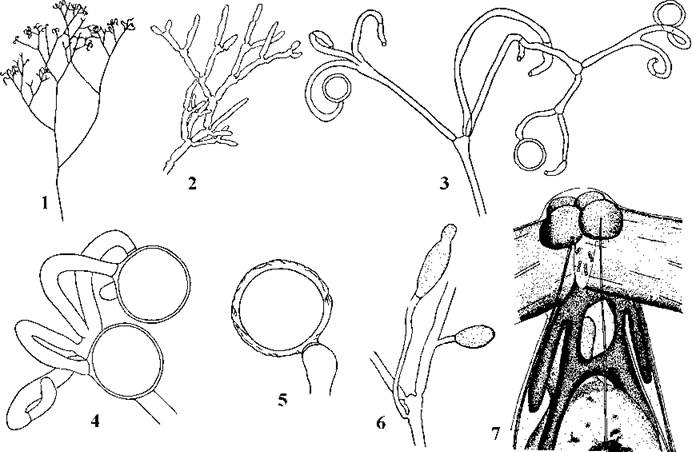
Рис. 24.56. Dichotomosiphon: 1 - зовнішній вигляд талому, 2 - фрагмент верхівкової частини талому, 3 - верхівковий сифон з оогоніями та антеридіями, 4 - зрілі оогонії та антеридій, 5 - зигота, 6 - фрагмент талому з вивідковими бруньками, 7 - апікальна частина сперматозоїду (1-6 - за Зауер, 1977; Starmach, 1972; 7 - за Moestrup, 1975).
За рядом ознак водорість подібна до галімедальних: фотосинтетичний апарат гетеропластидний, і включає як хлоропласти, так і амілопласти; у пластидах виявлено структури, що нагадують тилакоїд-утворюючі тіла, стефаноконтні стадії відсутні. Запасні полісахариди відкладаються не лише у стромі хлоропластів, але й безпосередньо у цитоплазмі (подібне явище спостерігається також у дазикладальних водоростей).
Водорість розмножується вивідковими бруньками та статевим шляхом. Антерідії та оогонії утворюються на верхівках бічних відгалужень талому. В антеридіях утворюється велика кількість дводжгутикових сперматозоїдів, в оогонії - одна яйцеклітина. Сперматозоїди виходять назовні, через невеличнкий сосочкоподібний носик проникають в оогоній та запліднюють яйцеклітину. Зигота одягається тришаруватою оболонкою, переходить у стан спокою, по закінченні якого проростає у новий сифональний дихотомічно розгалужений талом.
Dasycladales. Порядок являє собою самостійну чітко відокремлену еволюційну лінію сифональних водоростей з цілим комплексом специфічних ознак. Таломи мають радиально-симетричну будову, і являють собою вісь з системою апікальних мутовчастих сифонів. Клітинні оболонки складаються переважно з маннану, і просякнені карбонатами, найчастіше - у формі арагоніту. Фотосинтетичний апарат гомопластидний, представлений лише хлоропластами, що позбавлені піреноїдів і тилакоїд-організуючих тіл. Запасні полісахариди, подібно до Dichotomosiphon, відкладаються як у стромі хлоропласту, так і безпосередньо у цитоплазмі. Специфічною особливістю джгутикового апарату є відсутність на базальних тілах термінальних ковпачків. Первинне ядро відрізняється від ядер інших сифонофіцієвих наявністю перинуклеарних тіл.
Як і у інших сифонофіцієвих, в клітинах дазикладальних відбувається активний рух цитоплазми. Проте його специфічною особливістю є наявність двох потоків: відносно повільного поверхневого зі швидкістю біля 50-100 мкм/хв, та стрімкого глибинного, швидкість якого складає 200-600 мкм/хв. Рух цитоплазми у глибинному потоці відбувається не тільки за участю актину, але й тубуліну.
В онтогенезі дазикладальні проходять послідовно дво- або трьохрічну стадію одноядерної рослини з первинним ядром та короткочасну стадію власне сифональної рослини з багатьма вторинними ядрами. Статевий процес ізогамний, причому гамети утворюються не у гаметангіях, а в особливих гаметичних цистах.
Найбільш відомий представник порядку - рід Acetabularia. Таломи цієї водорості багаторічні. Протягом перших дво-трьох років вони складаються з центральної вісі 2-3 см завдовжки, ризоїдів та одного-трьох кілець незрослих між собою мутовок, розташованих на верхівці центральної вісі. При основі центральної вісі знаходиться одне велике первинне ядро, діаметр якого становить біля 100 мкм. Наприкінці вегетаційного сезону апікальна частина талому відмирає, і життєздатною лишається тільки ризоїдальна частина та невелика базальна ділянка центральної вісі з первинним ядром (рис. 24.57).
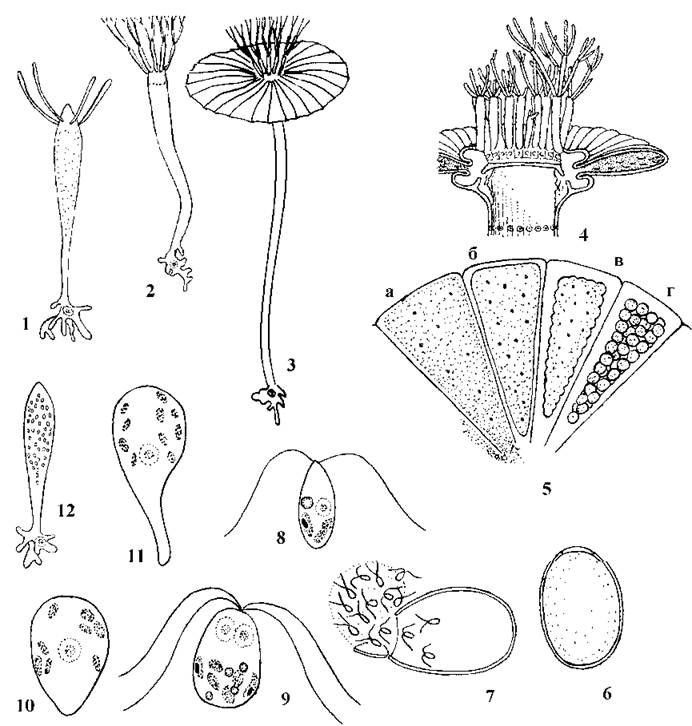
Рис. 24.57. Послідовні стадії циклу розвитку Acetabularia: 1, 2 - стерильні таломи першого та другого років з первинним ядром у ризоїдальній частині; 3 - фертильний талом третього року з сегментами, що утворюють парасольку; 4 - поперечний зріз через парасольку (2, 4 - за Виноградова, 1977; 1, 3, 5-12 - за "Entwicklung von Acetabularia (Dasycladales)", IWF, 1978).
На третьому-четвертому році життя апікальні мутовки зростаються між собою, утворюючи характерну для ацетабулярії "парасольку". В цей час первинне ядро у 2-3 рази зменшується у діаметрі, редукційно ділиться (соматична редукція), далі відбувається серія мітозів, внаслідок чого утворюється біля 20 000 дрібних гаплоїдних вторинних ядер. Ці ядра током цитоплазми виносяться в апікальну частину талому, зосереджуються у "парасольці", і водорість набуває справжньої сифональної будови. У променях парасольки протопласт водорості спочатку стискається, і далі розпадається на одноядерні ділянки. Останні одягаються товстою целюлозною оболонкою і перетворюються на гаметичні цисти з великим отвором, закритим спеціальною пробкою1.
Від кількох тижнів до кількох місяців цисти залишаються у "парасольці". Протягом цього періоду ядра повільно діляться і цисти стають багатоядерними. Далі "парасолька" руйнується, цисти звільняються, в них утворюється велика кількість одноядерних дводжгутикових ізогамет. Цисти відкриваються, гамети виходять назовні і копулюють. Зигота без періоду спокою одразу починає проростати у нову рослину з первинним ядром, ризоїдами, центральною вісью та кільцем незрослих апікальних мутовок. Таким чином, Acetabularia має диплогаплофазний життєвий цикл без зміни поколінь з соматичною редукцією.
Для роду Dasycladus характерні таломи, у яких центральна вісь вкрита багатьма близько розташованими кільцями мутовок. Кожна гілка мутовки тричі дихотомічно або трійчасто розгалужена (рис. 24.58).
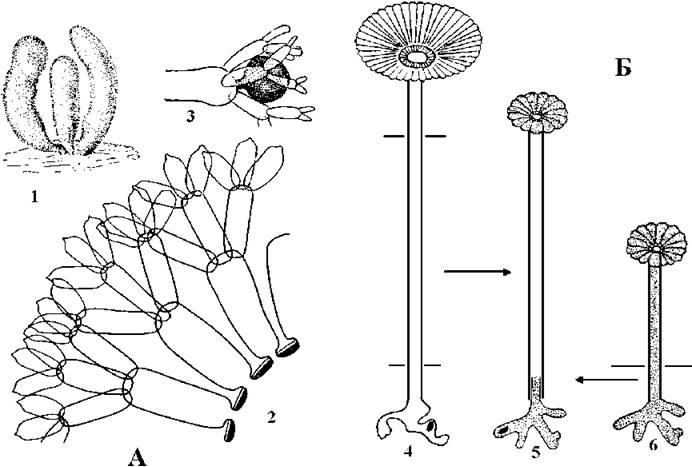
Рис. 24.58. Дазикладальні водорості. А - Dasycladus: 1 - зовнішній вигляд талому; 2 - фрагмент талому на поперечному зрізі; 3 - фертильний сферичний сегмент серед стерильних сегментів. Б - схема вегетативної гібридизації Acetabularia: позбавлену ядра ділянку "стебла" A.mediterranea (1, біла) прищеплювали на ділянку ризоїдальної системи з ядром A.wettsteinii (3, пунктирована). Трансплантована рослина (2) розвивала "парасольку" A.wettsteinii (А - за Виноградова, 1977; Б - за Масюк, 1993).
Всі дазикладальні водорості мешкають у морях, переважно тропчних та субтропічних. Так, види Acetabularia мешкають у теплих морях північної півкулі (зокрема, у Середземному морі поширеним видом є A.mediterranea). Інші дазикладальні є переважно мешканцями тропічних морів (наприклад, Dasycladus росте у Карибському морі). Завдяки наявності у клітинних оболонках великої кількості вапна, ці водорості добре зберігаються у викопному стані, що дало можливість прослідкувати напрямки еволюції даного порядку.
Найдавніші рештки Dasycladales відомі з верхнього докембрію протерозою (біля 1 млрд. років). За викопними рештками добре простежено морфологічну еволюцію в межах цього порядку: встановлено, що найпростіші дазикладальні водорості мали талом, позбавлений будь-яких асиміляційних пагонів, далі з'явились види з асиміляційними відростками, розміщеними на центральній вісі спочатку спірально, потім - супротивно і нарешті - мутовчасто, по всій довжині центральної вісі. У наймолодших форм мутовки зайняли лише апікальне положення.
Викопні дазикладальні відіграли помітну роль в утворенні карбонатних порід. Наприклад, деякі вапнякові масиви Альп утворені саме залишками таломів водоростей цього порядку. Завдяки наявності гігантського первинного ядра дазикладальні водорості стали популярним лабораторним об'єктом цитологічних та фізіологічних досліджень. Зокрема, саме на Acetabularia Хеммерлінгом були проведені класичні досліди по вивченню функції ядра та вегетативній гібридизації.
1 До недавнього часу вважалося, що мейоз відбувається саме на цій стадії, перед утворенням гамет. Проте сучасні електронно-мікроскопічні дослідження показали, що редукційний поділ має місце набагато раніше - при першому поділ первинного ядра.