Альгологія - Костіков І.Ю. - 2009-2013
Розділ 24. Зелені водорості – Chlorophyta
24.3. Стрептофітна лінія
Стрептофітна лінія об'єднує ті зелені водорості, які є близько спорідненими з вищими рослинами. Вважається, що саме до стрептофітної лінії належали пращури перших вищих рослин. До стрептофітів належать водорості, у яких цитокінез здійснюється за участю примітивного або розвиненого фрагмопласту, мітоз є відкритим або напіввідкритим; у представників, які мають джутикові стадій, цитоскелет монадних клітин є асиметричним. Лінію стрептофітів представляють два класи - Charophyceae та Zygnematophyceae (=Conjugatophyceae).
24.3.1. Клас Charophyceae
Харофіцієві водорості порявняно невеликі за обсягом. Клас об’єднує біля 400 видів сучасної флори та близько 1000 видів, відомих по викопних рештках. Проте Charophyceae є надзвичайно важливим у філогенетичному відношенні, оскільки саме примітивні харофіцієві стали пращурами вищих рослин.
Клас об’єднує мікро- та макроскопічні водорості, що, як і кон’югати, представляють фрагмопластну лінію еволюції. Проте, на відміну від Conjugatophyceae, харофіцієві зберегли джгутикові стадії, причому вони, подібно до сперматозоїдів моховидних та плауновидних, мають асиметричну систему джгутикових коренів та вкриті субмікроскопічними лусочками.
Представники класу зустрічаються у прісних та солоних водоймах, ґрунтах, аерофітоні.
Ознаки та характеристика класу
Біохімічні особливості
Склад пігментів та продуктів асиміляції у харофіцієвих такий само, як у кон’югат, хлорофіцієвих та требуксієфіцієвих водоростей, тобто специфічних біохімічних особливостей в межах класу не виявлено.
Цитологічні особливості
Клітинні покриви. Клітини харофіцієвих вкриті целюлозно-пектиновою оболонкою. У одного з порядків (Charales) синтез мікрофібрил целюлози здійснюється термінальними ферментними комплексами, а клітинні оболонки просякуються вапном. В інших порядках термінальні комплекси не виявлені, клітинні оболонки позбавлені карбонатів. Монадні клітини - зооспори та сперматозоїді - вкриті плазмалемою, на поверхні якої відкладаються субмікроскопічні органічні лусочки, подібні до лусочок монадних клітин кодіолальних та празинофіцієвих водоростей.
Ядерний апарат та особливості мітозу. Ядра мають одне-кілька ядерець, що, на відміну від кон’югат, на початку мітозу зникають. Багато специфічних особливостей виявлено в організації ядер харальних водоростей (див. нижче).
Мітоз відкритий. Центриолі є, і принаймі при утворенні монадних клітин, приймають участь у побудові веретена поділу. Веретно зберігається у телофазі, і стає центром організації фрагмопласту.
У кокоїдних, сарциноїдних та нитчастих форм, подібно до кон’югат, цитокінез відбувається внаслідок росту кільцевої борозни. Вростання кільцевої борозни в клітину обумовлено злиттям пухирців комплексу Гольджі з зачатковою борозною плазмалеми. Рух пухирців до борозни направляє система мікротрубочок залишків веретена та комплекс крайових мікротрубочок, що утворюється безпосередньо біля борозни поділу. Крім мікротрубочок, до складу систем, що керують рухом пухирців, входять мікрофібрили актину. Весь цей мікротубулярно-актиновий комплекс розглядається як фрагмопласт. Клітинна платівка та пори з плазмадесмами у кокоїдних, сарциноїдних та нитчастих форм не утворюються (рис. 24.59).
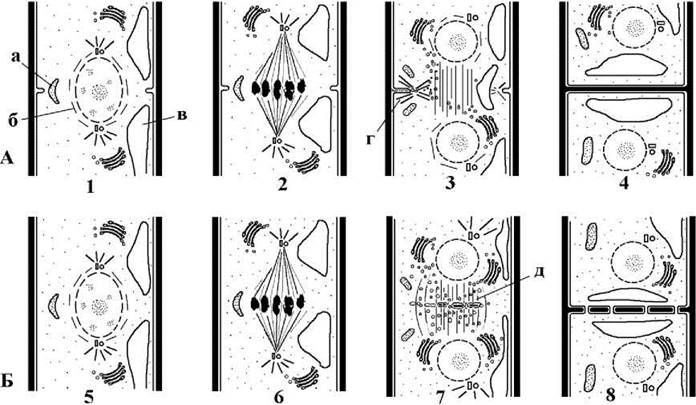
Рис. 24.59. Мітоз та цитокінез у харофіцієвих водоростей. А - поділ клітини у нитчастої водорості Klebsormidium; Б - поділ клітини у гетеротрихальної водорості Coleochaete; 1, 5 - профаза; 2, 6 - метафаза; 3, 7 - телофаза; 4, 8 - рання інтерфаза (а - мікротільце, б - перинуклеарні мікротрубочки, в - вакуоля з клітинним соком, г - крайові мікротрубочки фрагмопласту; д - фрагмопласт з клітинною платівкою) (А - за Lokhorst, Star, 1985; Б - за Marchant, Pickett-Heaps, 1973).
У гетеротрихальних представників цитокінез здійснюється тільки шляхом утворення клітинної платівки за участю фрагмопласту. Ріст платівки у всіх представників відбувається внаслідок злиття мембранних пухирців з матеріалом майбутньої клітинної перегородки, що відшнуровуються від комплексу Гольджі. Як результат формується клітинна платівка, що має пори з плазмадесмами.
Монадні стадії дводжгутикові, у високоспеціалізованих представників (Charales) представлені тільки сперматозоїдами, у інших - гаметами та зооспорами п-типу. Монадні клітини дорзовентральні, завжди вкриті одним шаром субмікроскопічних лусочок. Джгутики розміщуються на вентральному боці біля верхівки клітини і направлені не вперед, а в бік (т.з., унілатеральне положення).
Базальні тіла розходяться під невеликим кутом, і з’єднані між собою поперечносмугастою зв’язкою. Під базальними тілами починається один мікротубулярний корінь, що складається з кількох десятків мікротрубочок, розташованих в один ряд. Цей корінь під плазмалемою по вентральному боку клітини до її нижнього кінця. Безпосередньо біля базальних тіл мікротрубочки кореня переходять у багатошарову структуру (рис. 24.60). Подібний тип організації джгутикових коренів характерний для сперматозоїдів вищих рослин і не має аналогів в інших класах зелених водоростей.
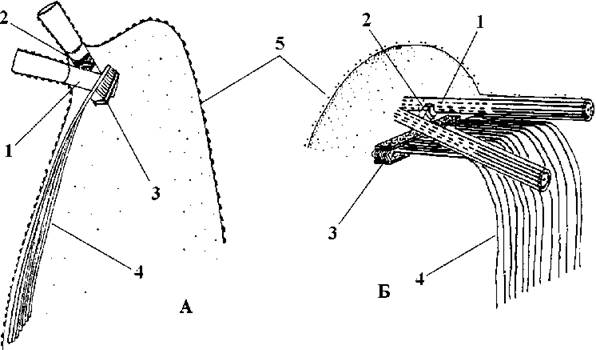
Рис. 24.60. Джгутиковий апарат сперматозоїдів Chara (А) та зооспор Chaetosphaeridium (Б): 1 - базальне тіло, 2 - верхне з"єднувальне волокно, 3 - багатошарова структура, 4 - унілатеральний мікротубулярний корінь з багатьох мікротрубочок (мікротрубочковий сплін), 5 - субмікроскопічні лусочки (А - за Масюк, 1989; Б - з Van den Hoek, Mann, Jahns, 1995 за Moestrup, 1982).
Типи морфологічних структур
В межах класу представлені водорості з чотирма типами морфологічних структур: кокоїдним, сарциноїдним, нитчастим та гетеротрихальним. Два перших типи сьогодні відомі лише у двох родів - Chaetoshpaeridium та Chlorokybus, відповідно. Два роди - Klebsormidium та Raphidonema - мають нитчастий тип структури і утворюють нерозгалужені однорядні нитки, які у ряду видів легко розпадаються на поодинокі клітини. Більшість харофіцієвих має гетеротрихальний тип структури. Таломи в цьому випадку можуть мати або вигляд сланких розгалужених ниток, що зростатючись утворюють паренхіматозні диски, від поверхні яких відходять безбарвні волоски (Coleochaete), або мати складну членисто-мутовчасту будову, нагадуючи хвощі (представники порядку Charales).
Розмноження та життєві цикли
Харофіцієві розмножуються як нестатевим, так і статевим шляхом. Нестатеве розмноження у неспеціалізованих форм відбувається поділом клітин надвоє, фрагментацією таломів, дводжгутиковими зооспорами та зрідка - акінетами. У високоспеціалізованих водоростей з порядку Charales поширено вегетативне розмноження вивідковими бруньками.
Статевий процес переважно оогамний, рідше - гетерогамний. Життєвий цикл у всіх еугамних харофіцієвих гаплофазний, з зиготичною редукцією, у кокоїдних та нитчастих форм - без зміни поколінь, у гетеротрихальних представників - з гетероморфною зміною. Агамні види нечисленні, представлені кокоїдними, сарциноїдними та кількома нитчастими формами; у них життєвий цикл являє собою простий цикломорфоз.
Система класу
За молекулярними критеріями, які добре узгоджуються з багатьма цитологічними та морфологічними ознаками, клас поділяється на п’ять порядків: Chaetoshpaeridales, Chlorokybales, Klebsormidiales, Coleochaetales та Charales. Основними фенотипічними ознаками порядків є тип морфологічної структури та план будови тіла (зокрема, наявність членисто-мутовчастої будови), особливості цитокінезу (зокрема, чи приймає в ньому участь кільцева борозна), здатність до розмноження за допомогою зооспор, наявність статевого процесу, та ін. (табл. 24.7).
Chaetoshpaeridales. До порядку відноситься лише один рід Chaetoshpaeridium з чотирма видами. За молекулярно-філогенетичними даними цей рід є найбільш примітивною групою харофіцієвих, яка майже одночасно дала початок всім іншим порядкам класу, а також кон’югатам та лінії, що безпосередньо привела до виникнення вищих рослин.
Табл. 24.7. Деякі основні систематичні ознаки порядків харофіцієвих водоростей
Ознака → Порядок ↓ |
Тип структури |
Чл.-мутовч. будова |
Зооспори |
Статевий процес |
Chaetoshpaeridales |
кокоїдний |
- |
+ |
оогамія |
Chlorokybales |
сарциноїдний |
- |
+ |
- |
Klebsormidiales |
нитчастий |
- |
+ |
гетеро-, оогамія |
Coleochaetales |
гетеротрихальний |
- |
+ |
оогамія |
Charales |
гетеротрихальний |
+ |
- |
оогамія |
Скорочення: Чл.-мутовч.будова - наявність членисто-мутовчастої будови талому (ознака стосується лише гетеротрихальних форм).
Види роду Chaetosphaeridium - це кокоїдні прісноводні перифітонні водорості. Клітини поодинокі або з’єднані спільним слизом чи слизовими тяжами у колонії. Кожна клітина, крім оболонки, вкрита слизовою капсулою та має довгу целюлозну щетинку, що є видозміненою частиною клітинної оболонки. У клітині міститься один-два парієтальних пластинчастих хлоропласта з піреноїдами та одне ядро. Протопласт обмежено рухливий - він разом з хлоропластами повільно обертається навколо вісі клітини (рис. 24.61).
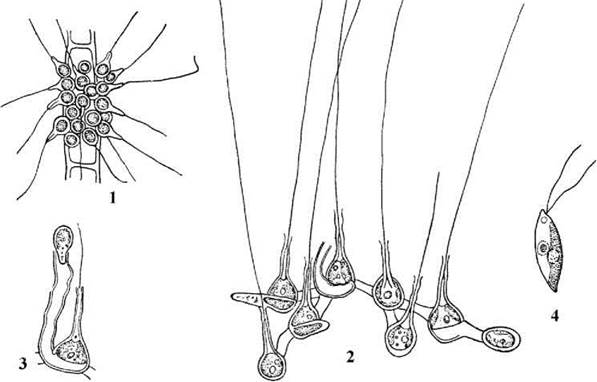
Рис. 24.61. Chaetosphaeridium: 1 - загальний вигляд колонії, 2 - утворення та вихід зооспор, 3 - з’єднання клітин у колонії, 4 - зооспора (1-3 - за Мошкова, 1979; 4 - ориг.).
Розмноження відбувається поділом клітини надвоє та зооспорами. У одного виду виявлено статевий процес. При поділі надвоє протопласт клітини ділиться у площині, паралельній субстрату, далі нижня клітина повертається на бік і продовжує рости, утворюючи невеликий циліндричний тяж, що прориває слизову обгортку материнської клітини. Далі у кожної дочірньої клітини утворюється власна клітинна оболонка, щетинка, нова слизова обгортка. Оболонка материнської клітини після цього ослизнюється. На жаль, електронно-мікроскопічні дані про особливості каріокінезу та цитокінезу поки що відсутні.
Зооспори дводжгутикові, дорзовентральні, позбавлені стигми і утворюються по чотири. Кожна зооспора вкрита плазмалемою, на поверхні якої є субмікроскопічні лусочки. Коренева система асиметрична, включає один мікротубулярний корінь, побудований приблизно з 60 мікротрубочок та багатошарову структуру.
Оогамний статевий процес виявлено C.globosum. Життєвий цикл у цієї водорості гаплофазний, з зиготичною редукцією та без зміни поколінь.
Chlorokybales. Порядок монотипний, і включає лише один вид - Chlorokybus atmophyticus, що мешкає у ґрунтах та на вологих аерофітних субстратах. Таломи водорості сарциноїдні, мають вигляд кубічних пакетів, одягнених загальним колоніальним слизом. Кожна клітина містить парієтальний лопатевий хлоропласт з голим піреноїдом. Між оболонкою та хлоропластом є особлива сферична структура, що отримала назву псевдопіреноїду. Цитокінез відбувається за участю кільцевої борозни, хоча у телофазі навколо веретена спостерігається утворення додаткових пучків мікротрубочок, що нагадують фрагмопласт (рис. 24.62).
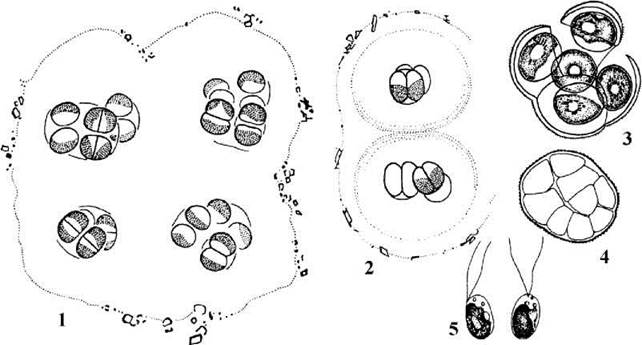
Рис. 24.62. Chlorokybus: 1, 2 - загальний вигляд колоній, 3 - апланоспорангій, 4 - зооспорангій, 5 - зооспори (за Geitler, 1955; Rith, 1972).
Розмноження відбувається фрагметацією пакетів, апланоспорами та дводжгутиковими зооспорами, що утворюються по одній з протопласту будь-якої клітини. Ультраструктура зооспор така ж сама, як у Chaetoshpaeridium.
Klebsormidiales. Об’єднує водорості з нитчастим типом структури. У клебсормідіальних цитокінез відбувається за участю кільцевої борозни, а поперечна клітинна перегородка позбавлена плазмадесм. Ультраструктура зооспор в цілому подібна до Chaetoshpaeridium. Як і у Chlorokybus, одна материнська клітина може дати початок лише одній монадній клітині - зооспорі або гаметі.
До порядку належить біля 40 видів двох родів - Klebsormidium та Raphidonema (рис. 24.63). Є дані про те, що деякі види роду Stichococcus з класу требуксієфіцієвих також належать до клебсормідіальних.
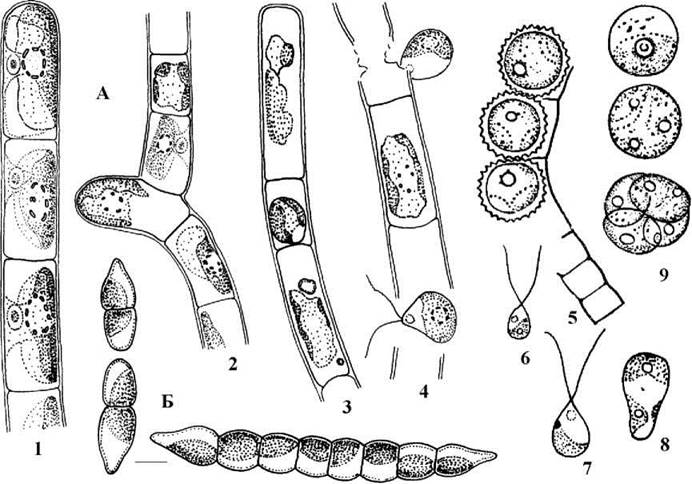
Рис. 24.63. Клебсормідіальні водорості. А - Klebsormidium (1 - гаметоспорофіт; 2 - зачаткове галуження; 3 - утворення зооспори; 4 - вихід зооспори; 5 - акінети; 6, 7 - мікро- та макрогамети; 8 - початкова стадія проростання зиготи; 9 - апланоспора та її проростання). Б - Raphidonema (А: 1-4, Б - ориг., 5-9 - за Мошкова, 1979).
Талом Klebsormidium має вигляд нерозгалуженої однорядної нитки, що зовні дуже подібна до Ulothrix. Клітини нитки морфологічно та функціонально однакові, кожна клітина містить один пристінний хлоропласт з піреноїдом. Розмножується Klebsormidium фрагментацією ниток, акінетами, зооспорами та статевим шляхами. Статевий процес гетерогамний. Гамети, крім того, здатні партеногенетично проростати у нові гаметоспорофіти, тобто виконувати функцію зооспор. Життєвий цикл гаплофазний, з зиготичною редукцією і без чергування поколінь
Види Klebsormidium надзвичайно широко поширені у ґрунтах (особливо степових та лучних), в аерофітоні, зрідка зустрічаються у прісних водоймах.
Raphidonema об’єднує переважно криофільні види, що мешкають на снігу та льодовиках. Нитки у цих водоростей короткі, складаються переважно лише з двох клітин і звичайно легко розпадаються. Самі клітини загострені на кінцях, містять один парієтальний хлоропласт з піреноїдом або без нього. Розмноження відбувається фрагментацією ниток. У одного виду - R.longiseta - відомий також оогамний статевий процес, причому сперматозоїди мають типову для клебсормідіальних дорзовентральну будову та асиметричний цитоскелет. Життєвий цикл гаплофазний, з зиготичною редукцією та без зміни поколінь.
Coleochaetales. До порядку входить один рід прісноводних гетеротрихальних водоростей - Coleochaete (біля 10 видів), таломи яких мають вигляд розгалужених сланких ниток або одношарових паренхіматозних платівок, як правило - з волосками (хетами) специфічної будови: кожний волосок при основі має комірець. Цитокінез відбувається шляхом утворення клітинної платівки за участю розвиненого фрагмопласту. На відміну від клебсормідіальних, поперечні перегородки мають пори з плазмадесмами, а кільцева борозна не приймає участі у поділі клітини. Подібно до хетосферидальних, протопласти клітин здатні до обмеженого осциляторного руху. У цитоплазмі міститься пристінний хлоропласт з одним-двома піреноїдами та одне ядро.
Розмножуються колеохетальні водорості фрагментацією таломів, голими дводжгутиковими зооспорами, подібними до зооспор Chaetosphaeridium, та статевим шляхом.
Статевий процес - оогамія. Яйцеклітина розвивається в оогонії, який у деяких видів має довгу шийку - трихогіну. В одноклітинних антеридіях утворюється по одному дводжгутиковому сперматозоїду. Цікаво, що сперматозоїди, як і у Klebsormidium, здатні проростати партеногенетично.
Після запліднення зигота переходить у стан спокою, при цьому від клітини з оогонієм та сусідніх клітин починають відростати гілочки, щільно притиснені до поверхні оогонію. Через це оогоній з заплідненою яйцеклітиною дещо нагадує архегоній примітивних вищих рослин. По закінченні періоду спокою у зиготі відбувається мейоз і зигота проростає у гаплоїдний спорофіт. Спорофіт має вигляд диску з 8-16 клиноподібних клітин. З кожної такої клітини утворюється по одній дводжгутиковій зооспорі, яка проростає у новий гаплоїдний гаметоспорофіт. Таким чином, життєвий цикл у Coleochaete гаплофазний, з зиготичною редукцією та гетероморфним чергуванням поколінь.
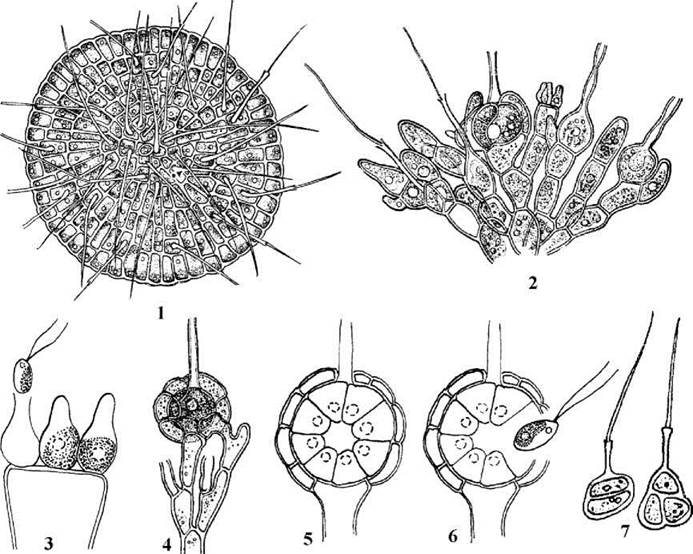
Рис. 24.64. Coleochaete: 1, 2 - зовнішній вигляд гаметоспорофіту у видів з таломами пластинчастої (C.scutata) та розгалужено-ниткоподібної (C.pulvinata) форми; 3 - антеридії та вихід сперматозоїду; 4 - зрілий оогоній, вкритий корою; 5 - восьмиклітинний спорофіт, що розвивається з зиготи; 6 - проростання спорофіту зооспорами; початкові стадії розвитку гаметоспорофіту (за Мошкова, 1979; Pringsheim, 1860).
Після відкриття подібності будови джгутикових стадій харофіцієвих водоростей та спорових вищих рослин порядок Coleochaetales розглядався як лінія, що дала початок мохоподібним і та судинним рослинам. Наявність гетероморфної зміни поколінь, гетеротрихальної будови зі здатністю формувати паренхіматозні диски, оогонію з трихогіною, архегоній-подібної будови оогоніїв з заплідненими яйцеклітинами суттєво підкріплювали це припущення. Проте молекулярно-філогенетичні реконструкції показали, що лінія, що призвела до виникнення вищих рослин відокремилась не від самих колеохетальних, а від їх пращура, який займав проміжне положення між Chaetosphaeridium, з одного боку, та Coleochaetales, Klebsormidiales та Chlorokybales, з іншого.
Charales. Об’єднує водорості, у яких таломи спорофітів нагадують хвощі: вони макроскопічні, складної членисто-мутовчастої будови і диференційовані на вузли та міжвузля (рис. 24.65). Представникам порядку притаманний цілий ряд специфічних цитологічних та морфологічних рис, життєвий цикл з гетероморфною зміною поколінь, причому гаметофіти ембріонізовані і розвиваються на спорофіті.
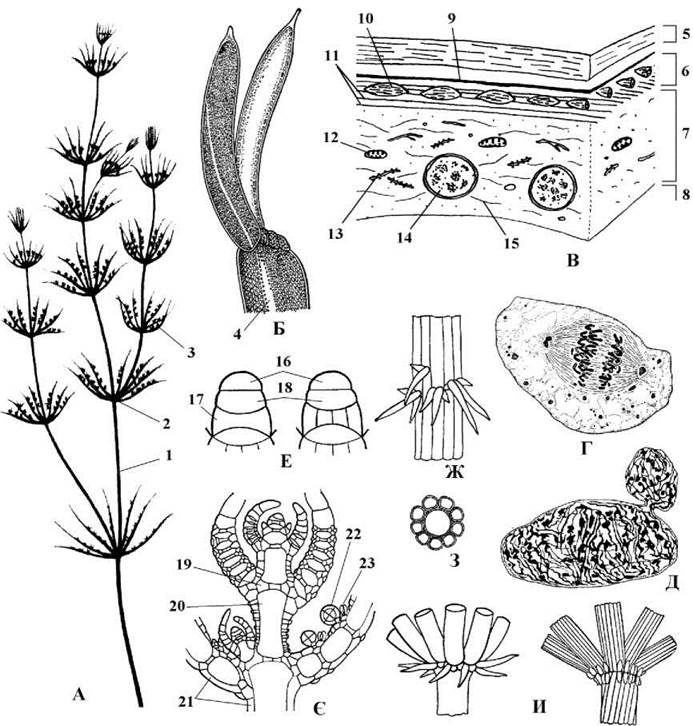
Рис. 24.65. Будова талому та особливості клітин спорофіту харальних водоростей. А - зовнішній вигляд талому Chara (1 - вузол, 2 - міжвузля, 3 - бічна "гілка" обмеженого росту); Б - верхівка гілки обмеженого росту Nitella (4 - лінія, що розділяє протилежні потоки цитоплазми); В - периферична частина клітини міжвузля (5 - клітинна оболонка, 6 - стабільна зона цитоплазми, 7 - лабільна зона цитоплазми, 8 - вакуоля з клітинним соком, 9 - плазмалема, 10 - хлоропласт, 11 - пучки мікрофібрил актину, що "направляють" потік цитоплазми, 12 - мітохондрія, 13 - ендоплазматична сітка, 14 - ядро, 15 - мікрофібрили міозину); Г - ядро клітини вузла в процесі мітозу; Д - ядро клітини міжвузля в процесі амітозу; Е - верхівкові клітини спорофіту (16 - апікальна клітина, 17 - подвійно-увігнута клітина, що дає початок клітинам вузла, 18 - подвійно-опукла клітина, що диференціюється у клітину міжвузля); Є - схема будови верхівки талому (19 - клітина вузла, 20 - клітина міжвузля, 21 - клітини стеблової кори, 22 - закладка антеридію, 23 - закладка оогонію); Ж - фрагмент міжвузля, вкритого стебловою корою з шипами; З - поперечний зріз через міжвузля зі стебловою корою; И - вузли з вінчиком "прилистників" (А-Б, Г-И - за Голлербах, 1977; В - за Stebbings, Hyams, 1979).
Мешкають харальні водорості у прісних та солоних континентальних водоймах та морських лиманах. До порядку входить біля 150 видів сучасної флори та більш 1000 викопних видів.
Цитологічні особливості. Клітини вкриті оболонками, в яких мікрофібрили целюлози синтезуються розетковими термінальними ферментними комплексами (подібний тип утворення оболонки характерний також для кон’югат та вищих рослин), і звичайно просякуються вапном.
Центр клітини зайнятий великою вакуолею з клітинним соком. Цитоплазма з органелами утворює тонкий периферичний шар. Вона диференційована на дві зони: стабільну та лабільну. Стабільна зона прилягає до плазмалеми і в ній одним шаром у правильних поздовжніх рядах розташовуються численні дрібні дископодібні хлоропласти, що позбавлені піреноїдів. Під стабільною зоною знаходиться лабільна зона. Тут зосереджені числені ядра, мітохондрії, елементи ендоплазматичної сітки та прикріплені до них мікрофібрили міозину; весь цей комплекс потоком цитоплазми швидко переміщується вздовж вісі клітини. Швидкість руху цитоплазми складає 50-100 мкм/секунду. Стабільна та лабільна зони розділені шаром паралельно розташованих пучків актину, що утворюють направляючі "рельси", по яких сковзуть мікрофібрили міозину разом зі з'єднаними з ними органелами. Потоки цитоплазми протилежних напрямків не змішуються між собою, і межа між ними має вигляд безбарвної смуги.
Клітини міжвузлів багатоядерні. Їх ядра мають характерну лопатеву форму, крім звичайних ядерних структур (хроматину, ядерець, прорибосом) вони містять пучки мікротрубочок і діляться амітотично. Клітини вузлів, навпаки, одноядерні, сферичні, їх поділ відбувається шляхом мітозу. Всі клітини спорофіту позбавлені центриолей. В клітинах чоловічих гаметофітів, навпаки, центриолі є і приймають участь у формуванні веретена поділу при утворенні сперматозоїдів. Мітоз відкритий. Цитокінез відбувається за участю фрагмопласту, причому у поперечних перегородках між дочірніми клітинами утворюються первинні пори з плазмадесмами.
Спорофіти харальних виглядають як розгалужені кущики, що складаються з прямостоячих опірних ниток необмеженого росту - т.з. "стебел", бічних мутовчасто розміщених асиміляторних ниток обмеженого росту - "листків", та ниток-ризоїдів. Кожна нитка ("стебло", "листок", ризоїд) диференційована на клітини вузлів та міжвузлів, що правильно чергуються одна з одною.
Вузли та міжвузля утворюються внаслідок поділу апікальної клітини. Вона ділиться у поперечній до вісі нитки площині, поперемінно відокремлюючи від себе клітини двох типів — подвійно-опуклу та подвійно-увігнуту. Подвійно-опукла клітина до подальшого поділу нездатна - вона лише росте та розтягується, поступово диференціюючись у велетенську багатоядерну клітину міжвузля. Подвійно-увігнута клітина здатна до подальших поділів. Ці поділи відбуваються у радіальних напрямках у поздовжній до осі клітини площині. Дочірні клітини утворюють вузол. Кожна клітина вузла, в свою чергу здатна, до поділу, який відбувається так само, як і в апікальній клітині, внаслідок чого утворюються "листки" та бічні гілки "стебла".
З клітин стеблових вузлів у деяких представників (наприклад, у Chara) розвиваються стеблова кора та прилистники, з листових вузлів - спорангії, що дають початок чоловічим або жіночим гаметофітам. На ризоїдальних вузлах утворюються вивідкові бруньки (бульбочки), що є органами вегетативного розмноження.
Гаметофіти розвиваються зі спорангіальних клітин, що утворюються у вузлах бічних гілок обмеженого росту. Жіночий гаметофіт складається з великої яйцеклітини та однієї або трьох дрібних супутніх клітин. Супутні клітини з'єднують гаметофіт зі спеціалізованою клітиною міжвузля спорофіту, яку називають клітиною-ніжкою. Від вузла, розташованого під клітиною-ніжкою відходять п’ять клітин кори оогонія. Вони по спіралі щільно оплітають гаметофіт і на верхівках відокремлюють одну або дві замикаючі клітини, що разом утворюють коронку. Відповідно, у першому випадку коронка одноярусна п’ятиклітинна, у другому - двоярусна десятиклітинна. Таким чином, дво- або чотирьохклітинний жіночий гаметофіт захищається десятьма або п'ятнадцятьма спеціализованими клітинами спорофіту. Система з гаметофіту, його корових клітин та коронки утворює оогоній (рис. 24.66).
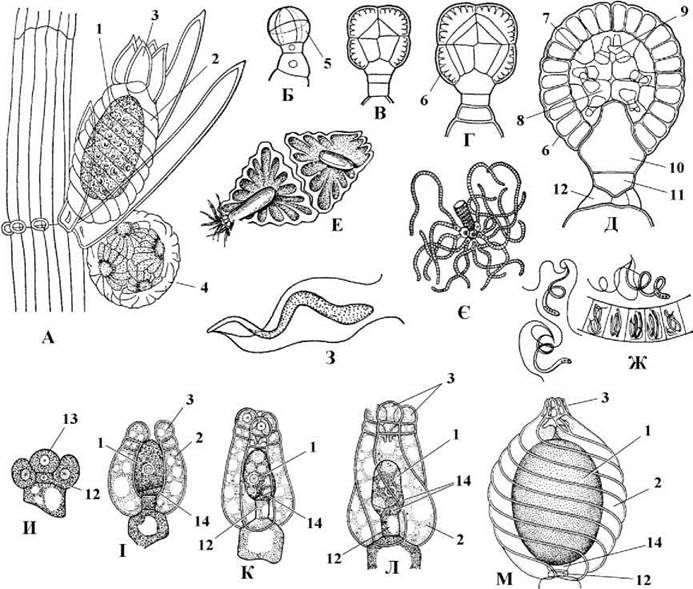
Рис. 24.66. Будова та розвиток гаметофіту харальних водоростей. А - фрагмент талому спорофіту Chara з жіночим та чоловічими гаметофітами; Б-Д - послідовні стадії розвитку антеридію; Е - октанти; Є - сперматогенні нитки; Ж - фрагмент сперматогенної нитки та вихід сперматозоїдів; З - сперматозоїд; И-М - послідовні стадії розвитку оогонію Nitella (1 - яйцеклітина, 2 - кора оогонію, 3 - коронка, 4 - чоловічі гаметофіти, що утворюють антеридій, 5 - октант, 6 - щиток, 7 - рукоятка, 8 - первинна головка, 9 - вторинна головка, 10, 11 - внутрішня (10) та зовнішня (11) базальні клітини, 12 - клітина листового вузла, 13 - ініціальна клітина жіночого гаметофіту, 14 - супутні клітини жіночого гаметофіту) (за Голлербах, 1977).
Чоловічі гаметофіти - октанти - багатоклітинні, вони з'єднуються по вісім, утворюючи сферичні антеридії. Октант складається з щитку, головок та сперматогенних ниток. Зовнішні клітини октантів - щитки - з'єднуються між собою боками, утворюючи замкнену сферу. Від щитка у порожнину сфери відходить одна клітина-рукоятка з однією первинною та шістьма вторинними клітинами-головками. Вторинні головки несуть на собі по чотири двохсотклітинні сперматогенні нитки, кожна клітина яких дає початок одному сперматозоїду. Сперматозоїди спірально скручені, мають два джгутики, асиметричну кореневу систему і вкриті субмікроскопічними лусочками.
Статевий процес оогамний. Зрілі щитки антеридію відокремлюються один від одного, оболонки клітин сперматогенних ниток ослизнюються і сперматозоїди потрапляють у воду. Далі вони підпливають до оогонію, клітини коронки при цьому роздвигаються, утворюючи отвір, через який сперматозоїд потрапляє до яйцеклітини. Після копуляції утворюється ребриста зигота - ооспора, яка переходить у стан спокою. По закінченні періоду спокою в зиготі відбувається мейоз, причому з чотирьох гаплоїдних ядер три дегенерують. Далі відбувається мітоз, одна з утворених клітин стає апікальною клітиною майбутнього стебла, друга - апікальною клітиною ризоїдів. Внаслідок поділу та подальшої диференціації клітин утворюється сланка (ризоїдальна) та висхідна частини нового талому (рис. 24.67). Таким чином, життєвий цикл харальних водоростей гаплофазний, з зиготичною редукцією, гетероморфною зміною поколінь та статевим диморфізмом.
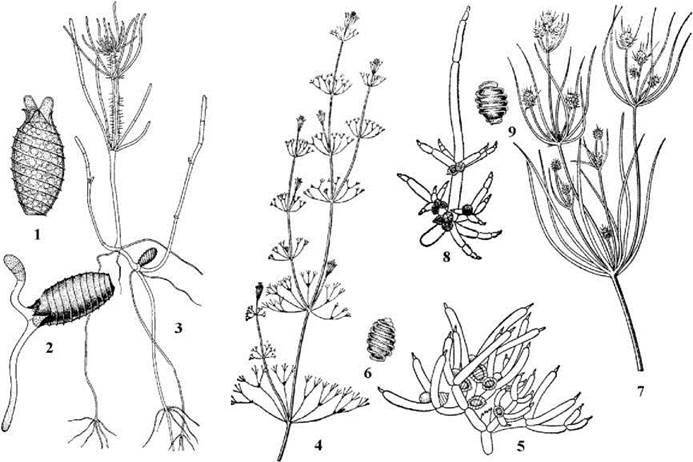
Рис. 24.67. Харальні водорості: 1-3 - проростання ооспори Chara; 4-6 - Nitella; 7-9 - Tolypella (1, 2 - ініціальні стадії розвитку ооспори; 3 - розвиток ризоїдальної та висхідної системи спорофіту; 4, 7 - спорофіти; 5, 8 - фрагменти талому з гаметофітами; 6, 9 - ооспори) (1-3 - за Голлербах, 1977; 4-9 - за Топачевський, Масюк, 1984).
Найбільш поширеними родами харальних водоростей є Chara, Nitella, Tolypella. Міжвузля у Chara вкриті стебловою корою, коронка оогоніїв п’ятиклітинна. У Nitella та Tolypella стеблова кора відсутня, а коронка складається з десяти клітин, розміщених у два яруси - по п’ять клітин у кожному. У водоймах ці водорості звичайно утворюють суцільні зарості і відіграють суттєву роль у продукційному режимі водойм. Ооспори (зиготи у стані спокою) є джерелом їжі для водоплаваючих птахів, особливо під час їх осінніх перельотів.
Викопні харальні водорості. Завдяки тому, що оболонки клітин, і особливо - кора оогонію накопичують велику кількість вапна, харальні добре зберігаються у викопному стані. Найдавніші рештки відомі з силуру (біля 420 млн. років тому). У девонському періоді харальні водорості досягли розквіту і були однією з пануючих груп мілинних ділянок світового океану. У відкладах цього періоду виявлено залишки більш, ніж 1000 видів. В кам'яновугільному періоді харальні водорості почали швидко зникати. У мезозойську еру (біля 200-100 млн. років тому) частина Charalesопанувала прісноводні біотопи, і біля 180 млн. років тому виникли перші сучасні види Charales.
В межах порядку за викопними рештками добре простежено еволюцію оогонію: від оогонію з порою через оогоній з коронкою у вигляді кришки до оогонію з коронкою та порою водночас. У палеозойських харальних водоростей кора оогонію була спірально завита зліва направо. У мезозої корові нитки оогонію розвернулися, і у сучасних форм оплітають яйцеклітину справа наліво.
Викопні харові водорості використовують при визначенні віку осадових порід у геології. Сучасні Charales є зручними об'єктами у фізіологічних та біофізичних дослідженнях. Зокрема, на прикладі клітин міжвузлів вивчаються процеси руху цитоплазми, електрофізіологічні явища (зокрема, генерацію поверхневих клітинних потенціалів та вплив їх на різні фізіологічні процеси у клітині).
Харофіцієві водорості та дискусії про місце харальних у системі рослинного світу
До 80-х років до Charophyceae відносили лише порядок Charales, і ознаками класу вважали перш за все наявність складної членисто-мутовчастой будови талому та специфічну будову статевих органів. Цим ознакам деякі провідні альгологи надавали настільки високого значення, що пропонували розглядати харальні водорості як самостійний відділ Charophyta (зокрема, ця точка зору відображена у виданні "Жизнь растений", довіднику "Водоросли", визначниках серій "Пресноводные водоросли СССР", "Визначник прісноводних водоростей Української РСР").
Прихильники системи Б. Фотта вважали харальні водорості самостійним класом в системі відділу зелених водоростей (наприклад, у короткому визначнику "Пресноводные водоросли Украинской ССР", підручнику "Algenkunde" та ін.). Результати порівняльно-цитологічних досліджень завершили дискусію "Представляють харальні водорості самостійний відділ чи є класом у системі Chlorophyta?" з висновком, що харальні водорості є лише одним з порядків класу харофіцієвих в межах відділу Chlorophyta.
Молекулярно-біологічні дані спочатку підтвердили цей висновок, оскільки Charales виявились спорідненими з Chlorokybales, Klebsormidiales та Coleochaetales. Проте в середині 90-х років дані сіквенсів ядерного гену, що кодує 18S РНК Chaetosphaeridium та різних кон’югат показали два неочікуваних результати: по-перше, кон’югати, вищі рослини та Chlorokybales-Klebsormidiales-Coleochaetales походять від нащадків Chaetosphaeridales і являють собою три паралельні еволюційні лінії; по-друге, Chaetosphaeridales та Charales є сестринськими таксонами, що походять від спільного пращура. Тому для збереження принципу монофілії треба або об"єднати кон’югати та харофіцієві в один клас, або розділити фрагмопластну лінію зелених водоростей на більшу кількість класів, зокрема, виділити в окремі класи по-перше, Chaetosphaeridales, по-друге, Charales, по-трете, Chlorokybales, Klebsormidiales та Coleochaetales. Тоді можливо зберегти кон’югати як клас.
Поширення та значення зелених водоростей у природі. Зелені водорості освоїли практично всі біотопи планети - моря та континентальні водойми, ґрунти та аерофітон, вони зустрічаються в гарячих джерелах та на снігу і льоді. У різних біотопах переважне поширення дістали різні класи зелених водоростей.
Характерними мешканцями морського планктону є представники класу Prasinophyceae. У морських бентосних та перифітонних біотопах серед зелених водоростей переважають види з Ulvophyceae та Siphonophyceae. Водорості з класів Chlorophyceae, Conjugatophyceae та Charophyceae опанували переважно прісноводні екотопи. В аерофітних угрупованнях зелені водорості представлені перш за все класом Trebouxiophyceae. Значна частина видів з Chlorophyceae, Trebouxiophyceae та Charophyceae мешкають також у ґрунтах.
Значення Chlorophyta у природі визначається, перш за все, тим, що зелені водорості, як і всі рослини, є першою ланкою трофічних ланцюгів. Біомаса зелених водоростей у водоймах може сягати значних величин: наприклад, при "цвітінні" води у гіпергалійних озерах біомаса Dunaliella salina інколи складає кілька кілограмів на м3, біомаса Chara у бентосних заростях або тини, що утворює Cladophora - до 3-5 кг/м2. В ґрунтах помірного поясу біомаса зелених водоростей в середньому становить 10-30 кг/га, а при "цвітінні" ґрунту - до 150-200 кг/га.
Річна продукція планктонних зелених водоростей у прісних мезотрофних водоймах (на 1 га) може сягати кількох центнерів, у бентосних морських угрупуваннях південних морів - навіть кількох тонн.
Зелені водорості є важливим компонентом, що обумовлює формування якості води та процеси її самоочищення. Серед усіх класів зелених водоростей знайдено представників, які здатні продукувати речовини з антибіотичною, інсектицидною, гербіцидною дією.
Одноклітинні види Chlorophyta нерідко викликають "цвітіння" води. Проте на відміну від "цвітіння", обумовленого масовим розвитком Cyanophyta або Dinophyta, воно, як правило, не шкідливе, а інколи (наприклад, у рибоводних ставках) є корисним. Деякі представники (Chloromonas, Chlamydomonas, Haematococcus, Raphidonema) є збудниками зеленого або червоного "цвітіння" снігу, і суттєво впливають на гляциологічний режим у горах. Вольвокальні, хлорококальні, клебсормідіальні водорості здатні викликати "цвітіння" ґрунту, а роди Desmococcus, Stichococcus, Trentepohlia- "цвітіння" кори дерев та поверхні різних надземних споруд.
В межах відділу значна частина видів живе у симбіозі з різноманітними тваринами, грибами та вищими рослинами. Зокрема, серед дрібних празинофіцієвих водоростей є ендосимбіонти радіолярій та гетеротрофної динофітової водорості Noctiluca. Tetraselmis convolutae мешкає у клітинах тубелярії Convoluta roscoffensis. Кілька видів роду Chlorella є фотоавтотрофними симбіонтами інфузорій та губок. Значна кількість зелених водоростей, переважно - з класу Trebouxiophyceae, є фікобіонтами лишайників (наприклад, Trebouxia, Trentepohlia).
Серед Chlorophyta відомі і деякі паразитичні представники. Зокрема, Prototheca паразитує на хребетних (в тому числі, і людині), викликаючи захворювання прототекоз. На вищих рослинах та водоростях найчастіше паразитують ульвофіцієві. Так, Cephaleuros спричинює захворювання листків чагарників та дерев у тропіках та субтропіках, деякі види Chlorochytrium паразитують у міжклітинниках ряски, Chlorocystis та стадія Codiolum є паразитами морських зелених та червоних водоростей-макрофітів.
Зелені водорості разом з синьозеленими є піонерами заростання новоутворених субстратів (наприклад, вулканічних островів) та першою ланкою у процесах вивітрювання гірських порід, на поверхні яких вони оселяються навіть раніш від лишайників.
Значення в господарській діяльності людини. Зелені водорості займають помітне місце у господарській діяльності людини. Вони є об’єктами фікотехнологічних виробництв та аквакультури, індикаторами стану оточуючого середовища, маркерами віку геологічних порід.
Найбільш відомі фікотехнологічні об’єкти - Dunaliella salina, Chlorella vulgaris, види Scenedesmus та Chlamydomonas. Гіпергалійна водорость Dunaliella salina вирощується як джерело Р-каротину, вміст якого при певних умовах складає біля 2% від сухої ваги клітини. Крім того, з Dunaliella отримують гліцерин, вуглеводневі сполуки, харчові барвники, виготовляють радіопротекторні та антиоксидантні препарати.
Chlorella vulgaris була першою водорістю, яка стала об’єктом інтенсивного промислового культивування. Хлорела використовується як вітамінні добавка та дієтичний продукт у раціоні людини, біостимулятор у тваринництві. Вона також інтенсивно вивчалась у космічних умовах з метою створення замкнених екосистем. Проте в цілому ця водорість виявилась невдалим біотехнологічним об’єктом, оскільки її клітини мають дуже міцну та хімічно стійку оболонку з шаром спорополеніну. Вдалішим об’єктом, придатним до використання з тією ж метою, що і Chlorella, виявився Scenedesmus.
Виділені з ґрунтів види роду Chlamydomonas знайшли використання у сільському господарстві як ґрунтозахисний та водоутримуючий засіб. Це пов’язано з тим, що підібрані з технологічною метою хламідомонади виробляють у ґрунті значну кількість слизу (у пальмелеподібному стані), у якій запасається волога та завдяки якій агрегуються ґрунтові частки. Найбільш відомим об’єктом у цій галузі став Chlamydomonas mexicana.
Їстівні ульвофіцієві водорості Ulva, Enteromorpha, Monostroma - т.з. зелені морські салати - є об’єктами морської аквакультури і в теплих морях вирощуються на морських фермах. Крім того, збір цих водоростей проводять і безпосередньо у природі.
Численна група зелених водоростей є індикаторами стану навколишнього середовища. Деякі види увійшли в атласи сапробних організмів і використовуються при розрахунках індексів сапробності (найбільшого визнання дістав метод Пантле та Бука). Аерофітна водорість Desmococcus vulgaris стала одним з тест-об’єктів на забруднення повітря, і тест за десмококом входить до переліку тестів міжнародної системи моніторингу навколишнього середовища.
В цілому, галузі використання зелених водоростей дуже різноманітні, і кількість патентних документів по вирощуванню та використанню Chlorophyta наприкінці ХХ ст. перевищила три тисячі.