Альгологія - Костіков І.Ю. - 2009-2013
Розділ 24. Зелені водорості – Chlorophyta
24.3. Стрептофітна лінія
24.3.2. Клас Zygnematophyceae (=Conjugatophyceae)
Кон’югати є одним з найчисленніших класів Chlorophyta, і нараховують біля 10 000 видів. Представники класу - переважно мікроскопічні водорості, мешканці прісних водойм. Дуже невелика кількість видів кон’югат зустрічається у солоних водоймах та у позаводних умовах (у ґрунтах та в аерофітоні). Кон’югати, разом з харофіцієвими водоростями, представляють фрагмопластну лінію еволюції зелених водоростей, яка дала початок вищим наземним рослинам. Специфічними рисами класу є повна відсутність будь-яких монадних стадій та наявність статевого процесу типу кон’югації.
Ознаки та характеристика класу
Біохімічні особливості
Склад пігментів та продуктів асиміляції є типовим для зелених водоростей в цілому (хлорофіли "а" та "в", всі типи каротинів, ксантофіли лютеінового ряду, крохмаль як основний продукт асиміляції), і таким чином, специфічні біохімічні особливості в межах класу відсутні.
Цитологічні особливості
Клітинні покриви представлені целюлозно-пектиновою оболонкою, причому синтез мікрофібрил целюлози відбувається внаслідок роботи розеткових термінальних ферментних комплексів, розташованих у плазмалемі. Кожний такий комплекс складається з шістьох субодиниць целлюлоза-синтетази, розміщених у вигляді розетки.
В цілому, клітинна оболонка кон’югат тришарова: зовні розташовується пектиновий шар, здатний утворювати масивний слиз. Проміжний та внутрішній шари є целюлозними. Мікрофібрили целюлози у проміжному шарі розташовані у вигляді безладно орієнтованих пучків. Целюлозна мікрофібрили внутрішнього шару утворюють правильні паралельні ряди.
У багатьох одноклітинних представників в клітинних оболонках є великі або дрібні пори. Через перші секретується слиз, обумовлюючи здатність клітин до повільного руху. Тому великі пори інколи називають також слизовими. Через дрібні пори відбувається обмін речовинами між клітиною та середовищем. У десмідіальних та гонатозигальних водоростей клітинні оболонки складно скульптуровані - вони мають різноманітні потовщення, вирости, шипики, щетинки.
Ядерний апарат та особливості мітозу. Ядра кон’югат звичайно добре помітні в оптичний мікроскоп завдяки наявності великого ядерця. На початку мітозу ядерце не зникає, а ділиться як самостійна органела, через що його інколи називають ендосомою.
Мітоз у кон’югат напіввідкритий, центріолі відсутні, веретено зберігається протягом всієї телофази. Цікаво, що ядерна оболонка зберігається протягом профази, метафази та анафази, хоча в ній або утворюються великі "вікна", або вона частково фрагментується. У телофазі ядерна оболонка зникає, і відновлюється лише по закінченні каріокінезу внаслідок злиття мембранних пухирців, що походять від комплексу Гольджі та ендоплазматичної сітки.
Хромосоми у кон’югат звичайно дрібні та численні, а положення центромерів не фіксоване. У багатьох видів кількість хромосом навіть в межах одного клону не постійна.
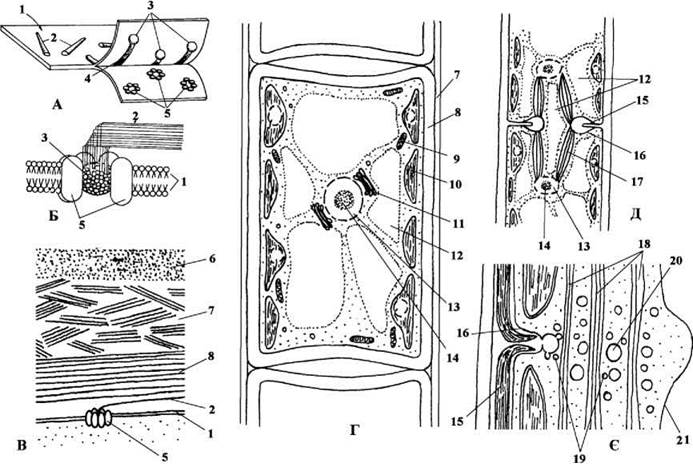
Рис. 24.68. Деякі ознаки класу Zygnematophyceae (=Conjugatophyceae). А, Б - схема будови розеткового термінального ферментного комплексу; В - будова клітинної оболонки; Г - загальний план будови клітини Д - телофаза мітозу; Є - утворення поперечної клітинної перегородки в телофазі (Г-Є - на прикладі Spirogyra);. 1 - плазмалема, 2 - мікрофібрила целюлози, утворена розетковим комплексом, 3 - центральна глобула целюлоза-синтетазного комплексу; 4 - відбиток мікрофибрили целюлози, 5 - розетка целюлоза-синтетазного комплексу, 6 - пектиновий шар оболонки, 7 - проміжний шар оболонки з невпорядковано розташованих мікрофібрил целюлози, 8 - внутрішній шар оболонки з впорядкованими мікрофібрилами целюлози; 9 - мітохондріальний профіль, 10 - хлоропласт, 11 - комплекс Гольджі, 12 - тонопласт, 13 - ядро, 14 - ендосома (ядерце), 15 - поперечна перегородка в процесі утворення, 16 - плазмалема в зоні борозни поділу, 17 - веретено поділу, 18 - мікротрубочки фрагмопласту, 19, 20 - дрібні та великі мембранні пухирці з матеріалом майбутньої поперечної перегородки, 21 -мембрана тонопласту (А-В - за Giddings, Brower, Staehelin, 1980; Кордюм, 1989; Mix, Manshard, 1977; Г - ориг., Д, Є - Fowke, Pickett-Heaps, 1969).
Цитокінез відбувається внаслідок одночасного росту кільцевої борозни та клітинної платівки. Остання утворюється за участю фрагмопласту: у телофазі відбувається збірка нових мікротрубочок та мікрофібрил актину, що орієнтуються паралельно до мікротрубочок веретена. Від комплексу Гольджі відшнуровуються пухирці, що містять матеріал майбутньої клітинної перегородки, і переміщуються в площину поділу клітини. Напрямком руху пухирців "керують" мікротрубочки та мікрофіламенти фрагмопласту. Клітинна платівка утворюється та росте внаслідок злиття цих пухирців, причому в ній немає пор, і тому дочірні клітини не з'єднуються між собою плазмадесмами.
Особливості фотосинтетичного апарату. Серед кон’югат переважають види з центральними хлоропластами зірчастої або пластинчастої форми, або з пристінними хлоропластами у вигляді спіралеподібних стрічок. Сітчасті, дископодібні та чашоподібні парієтальні хлоропласти, на відміну від інших класів, для кон’югат не характерні.
Монадні стадії повністю відсутні.
Типи морфологічних структур
В межах класу представлено лише два типи морфологічних структур: кокоїдний та нитчастий. У кокоїдних форм клітини поодинокі або об'єднуються у ланцюгоподібні колонії. Клітини десмідіальних водоростей посередині перетягнуті, і складаються із двох напівклітин. У мезотеніальних водоростей клітини циліндричні, прямі або зігнуті і на напівклітини не диференційовані.
Нитчасті форми мають вигляд довгих чи коротких однорядних нерозгалужених ниток. Інколи нитки легко розпадаються на поодинокі клітини.
Розмноження та життєві цикли
Розмноження відбувається вегетативним та статевим шляхами. Основним способом вегетативного розмноження одноклітинних форм є поділ клітин навпіл, у нитчастих - фрагментація ниток; деякі види здатні утворювати акінети. Статевий процес - кон’югація.
Життєвий цикл гаплофазний, без чергування поколінь, з зиготичною редукцією. Зигота проростає після періоду спокою 1-4 проростками.
Система класу
За типами структури, наявністю диференціації клітин на напівклітини, скульптурованістю оболонок, наявністю порового апарату, кількістю проростків у зиготи Conjugatophyceae за класичною системою поділяють на 4 порядки: Mesotaeniales, Desmidiales, Gonatozygales, Zygnematales.
Проте цей поділ лише частково співпадає з результатами молекулярно- філогенетичних реконструкцій, зокрема, за ядерним геном, що кодує 18S субодиницю рибосомальної РНК. Так, згідно до молекулярного дерева, клас включає три еволюційні лінії: 1) кокоїдні та нитчасті водорості з гладкою клітинною оболонкою без порового апарату, причому клітини у кокоїдних форм не диференційовані на напівклітини. Лінія відповідає монофілетичному порядку Mesotaeniales та парафілетичному порядку Zygnematales; 2) кокоїдні водорості зі скульптурованими оболонками або оболонками, що мають поровий апарат, проте не диференційовані на напівклітини. Лінія відповідає об'єднує порядок Gonatozygales та родини Peniaceae та Closteriaceae, що за класичною системою належать до Desmidiales; 3) кокоїдні водорості, клітини яких диференційовані на напівклітини та мають скульптуровані оболонки з розвиненим поровим апаратом. Лінія відповідає родині Desmidiaceae однойменного порядку. Фенотипічни ознаки порядків подані у таблиці 24.8.
Табл. 24.8. Основні систематичні ознаки порядків кон’югатофіцієвих водоростей, що корелюють з молекулярно-філогенетичними реконструкціями
Ознака → Порядок ↓ |
Напівклітини |
Оболонка: (скульптурованість або навність пор) |
Тип структури |
Zygnematales |
- |
- |
кокоїдний або нитчастий |
Gonatozygales |
- |
+ |
кокоїдний |
Desmidiales |
+ |
+ |
кокоїдний |
Zygnematales включає біля 1600 видів кокоїдних та нитчастих кон’югат. Клітини зигнематальних водоростей циліндричні або веретеноподібні, з гладкими оболонками позбавленими порового апарату. Через це зигнематальні, на відміну від інших порядків, не здатні до активного руху. У нитчастих форм нитки прості, однорядні, нерозгалужені і звичайно вкриті товстим шаром аморфного слизу. У водоймах ці водорості утворюють м’яке та слизьке на дотик ватоподібне жабуриння.
Розмноження відбувається поперечним поділом клітин навпіл, фрагментацією ниток та внаслідок кон’югації (рис. 24.69). При кон"югації у кокоїдних форм дві клітини одягаються спільним слизом, далі від кожної клітини у напрямку до іншої відростає короткий копуляційний відросток. У місці контакту копуляційних відростків клітинна оболонка розчиняється, протопласти клітин зливаються і утворюється зигота.
У нитчастих зигнематальних розрізняють два типи кон’югації - драбинчасту та бічну. При драбинчастій кон’югації зливаються протопласти клітин різних ниток: дві нитки одягаються спільним слизом, від кожної клітини нитки у напрямку до клітин нитки - партнера починає рости копуляційний відросток. Коли копуляційні відростки стикаються, оболонки у зоні контакту розчиняються і утворюється кон’югаційний канал. Далі або обидва протопласта рухаються назустріч один до одного і зливаються у кон’югаційному каналі (цей процес розглядається як фізіологічна ізогамія), або протопласт однієї клітини перетікає в іншу, і тоді зигота утворюється в приймаючий клітині (фізіологічна анізогамія). Драбинчастою таку кон’югацію називають через подібність двох ниток зі зрослими копуляційними відростками до драбини. Бічна кон’югація відбувається між двома сусідніми клітинами однієї нитки (рис. 24.69).
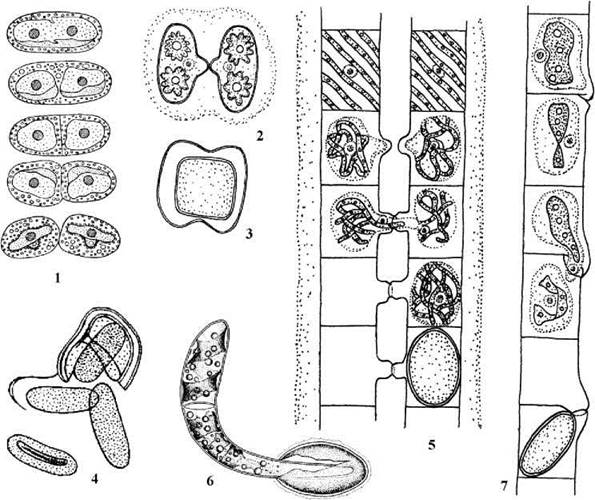
Рис. 24.69. Розмноження у зигнематальних водоростей. 1 - послідовні стадії поділу клітини Mesotaenium; 2, 3 — кон’югація (2) та зигота (3) у Cylindrocystis; 5, 6 - драбинчаста кон’югація (5) та проростання зиготи (6) у Spirogyra; 7 - бічна кон’югація у Mougeotia (14, 6 - за Рундина, 1977; 5, 7 - ориг.).
Зигота як у кокоїдних, так і у нитчастих форм одягається тришаровою товстою, звичайно - орнаментованою оболонкою і перетворюється на зигоспору, яка переходить у стан спокою. По закінченні періоду спокою ядро редукційно ділиться. У одноклітинних форм звичайно навколо кожного з чотирьох гаплоїдних ядер відособлюється частина цитоплазми, формується власна клітинна оболонка, і таким чином, утворюється чотири нових вегетативних клітини. У багатьох нитчастих видів три гаплоїдних ядра дегенерують, і тоді зигота проростає лише одним проростком. Тенденція зменшення кількості життєздатних ядер після мейозу у кон’югат розглядається як прогресивне явище, оскільки життєздатний проросток отримує більшу кількість поживних речовин, необхідних для його розвитку.
Рання дивергенція в межах порядку, за молекулярними даними, відбувалась за морфологічними типами пластид. Це призвело до утворення трьох еволюційних груп таксонів - а) з 1-2 центральними зірчастими хлоропластами; б) з осьовим пластинчастим хлоропластом; в) з пристінними спіралеподібними хлоропластами. Кожна з цих ліній дала початок нитчастим формам з відповідним типом хлоропласту, які за класичною системою вважали самостійним порядком Zygnematales, тоді як кокоїдні форми об’єднували у порядок Mesotaeniales.
Лінію водоростей з осьовим пластинчастим хлоропластом ілюструють кокоїдна водорість Mesotaenium та нитчаста водорість Mougeotia. Види роду Mesotaenium є мешканцями переважно вологих скель та епіфітами мохів та лишайників. Клітини циліндричні, поодинокі, з одним хлоропластом у вигляді осьової платівки. Від цього роду безпосередньо виводяться нитчасті водорості роду Mougeotia.
Талом Mougeotia має вигляд довгих або коротких нерозгалужених ниток, які у багатьох видів легко розпадаються на поодинокі клітини. Хлоропласт, подібно до Mesotaenium, має вигляд осьовьої платівки з численними піреноїдами. Цікаво, що в обох родах хлоропласт здатний змінювати своє положення залежно від інтенсивності освітлення. Так, при слабкому освітленні він повертається "анфас", перпендикулярно до падаючих променів, при інтенсивному - навпаки, у профіль, у площину паралельну до джерела світла.
Прикладами кокоїдних та нитчастих зигнематальних, у яких клітини містять 1-2 зірчасті хлоропласти, є Cylindrocystis та Zygnema, відповідно. Перший рід об’єднує переважно мешканців вологих ґрунтів, другий - прісноводні форми.
Лінію водоростей зі спіральними стрічкоподібними хлоропластами представляють кокоїдна Spirotaenia та нитчаста Spirogyra. У Spirotaenia клітини поодинокі, циліндричні, з одним-кількома пристінними стрічкоподібними спірально закрученими хлоропластами.
Представники цього роду найчастіше зустрічаються у болотах та на вологих позаводних субстратах.
Рід близько споріднений з прісноводною нитчастою водорістю Spirogyra, для якої також характерні хлоропласти у вигляді однієї або кількох спіралеподібних стрічок. Клітини спірогіри досить великі, їх більшу частину займає вакуоля з клітинним соком. В центрі вакуолі знаходиться невелика ділянка цитоплазми з добре помітним ядром. Ця ділянка з’єднується з периферичною частиною цитоплазми за допомогою кількох цитоплазматичних тяжів.
Одноклітинні зигнематальні водорості зустрічаються переважно у вологих аерофітних екотопах, а нитчасті - у прісних континентальних водоймах, звичайно - стоячих або з уповільненою течією, де зазвичай утворюють м’які слизькі тини. З такої тини у невеликих кількостях виготовляють високоякісний папір; на півночі висушені тини зигнематальних водоростей використовують як утеплюючий матеріал.
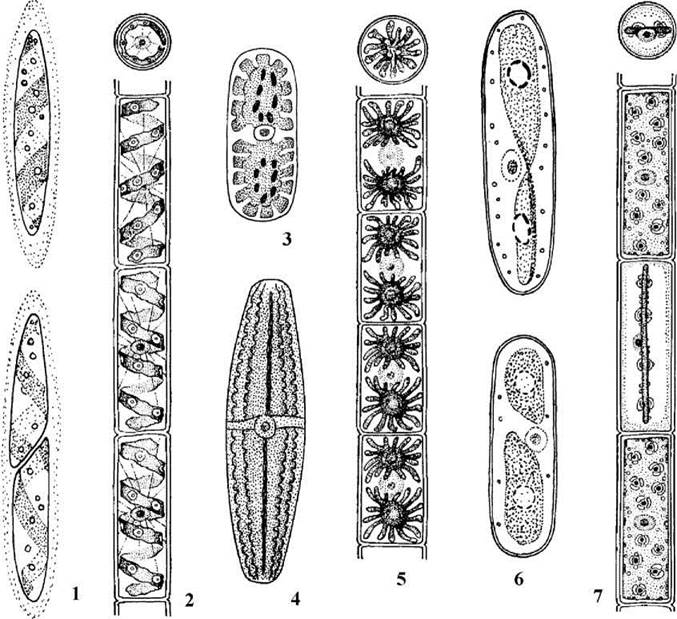
Рис. 24.70. Одно- та багатоклітинні представники Zygnematales: 1 - Spirotaenia, 2 - Spirogyra, 3 - Cylindrocystis, 4 - Netrium, 5 - Zygnema, 6 - Mesotaenium, 7 - Mougeotia (зверху у позиціях 2, 5, 7 показано поперечні зрізи через клітину) (1, 2, 5, 7 - за Рундина, 1977; 3, 4 - за Топачевский, Масюк, 1984; 6 - ориг.).
Gonatozygales. Нараховує біля 100 видів, переважна більшість яких належить до роду Closterium. Порядок об’єднує кокоїдні одноклітинні та колоніальні водорості, клітини яких не диференційовани на напівклітини, проте мають або розвинений поровий апарат, або скульптуровані оболонки, звичайно - хвилястих обрисів, або як перше, так і друге разом. Зигота проростає одним чи двома проростками. В межах порядку представлено три еволюційні лінії, які відповідають трьом родинам - Peniaceae, Gonatozygaceae та Closteriaceae.
Найбільш примітивним представником порядку, який дещо нагадує мезотеніальні водорості, є рід Penium (рис. 24.71). Водорість має циліндричні клітини, вкриті оболонкою, поверхня якої скульптурована дрібними гранулами або поздовжніми чи спіральними штрихами і позбавлена пор. Хлоропласт осьовий, з кількома поздовжніми ребрами та кількома піреноїдами, звичайно злегка перетягнутий посередині. На полюсах клітини інколи спостерігаються вакуолі, всередині яких знаходяться дрібні кристали гіпсу. Види роду мешкають зазвичай на вологих скелях та каміннях та на епіфітують на мохах.
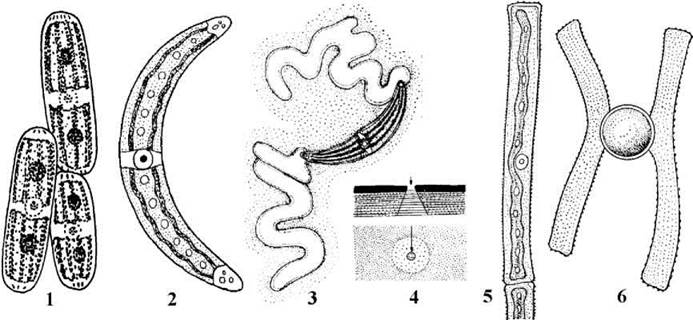
Рис. 24.71. Гонатозигальні водорості: 1 - Penium; 2 - 4 - Closterium (2 - вегетативна клітина, 3 - виділення слизу, препарат забарвлений розчином туші, 4 - будова пори на зрізі та при вигляді зверху); 5, 6 - Gonatozygon (5 - вегетативна клітина, 6 - зигота) (1 - ориг., 2, 5, 6 - за Топачевський, Масюк, 1984; 3, 4 - за Паламар-Мордвинцева, 1989).
У роду Closterium клітини також поодинокі, веретеноподібно зігнуті. Клітинна оболонка гладка або злегка штрихувата і має багато дрібних та дві великі пори. Останні знаходяться на полюсах клітини і виконують функції локомоторних органів - через них клітина виділяє слиз. Виділення слизу або спричинює повільний реактивний рух, або слиз утворює швидко ростучі слизові ніжки, за допомогою якого клітина змінює своє положення у просторі. При слабкому освітленні у клітин спостерігається позитивний фототаксис, при інтенсивному - негативний. Хлоропласти нагадують осьові конуси з радіальними поздовжніми ребрами; по осі хлоропласту розташовується кілька піреноїдів. Як і у Penium, на полюсах клітин утворюються вакуолі з кристалами гіпсу. У обох родів зигота проростає двома проростками.
Типовий рід порядку - Gonatozygon - має циліндричні клітини, які, з'єднуючись кінцями, утворюють малоклітинні нитки, що легко розпадаються на поодинокі клітини. Оболонки звичайно скульптуровані дрібними зернятками або шипиками і мають пори. Хлоропласти осьові, з піреноїдами. На кінцях клітин інколи спостерігаються вакуолі з кристалами гіпсу, подібні до вакуолей Closterium. Зигота звичайно проростає одним проростком. Види роду часто зустрічаються у різноманітних прісних водоймах, особливо навесні.
Desmidiales. Включає за різними оцінками 4-8 тис. видів кокоїдних водоростей. Клітини десмідіальних водоростей поодинокі або з'єднуються у стрічкоподібні колонії. Кожна клітина складається з двох симетричних напівклітин, що з'єднуються між собою широким або вузьким перешийком. Кут між напівклітинами називають синусом. Клітинні оболонки десмідіальних водоростей звичайно скульптуровані бородавками, шипами, шипиками, нерідко просякуються солями заліза, мають складний поровий апарат, що складається з багатьох дрібних та двох-чотирьох великих слизових пор, розташованих на полюсах клітин. Слизові пори забезпечують рух клітини, механізм якого такий самий, як і у Closterium. Кожна напівклітина містить один або два хлоропласти з піреноїдами, а в зоні перешийку - одне ядро.
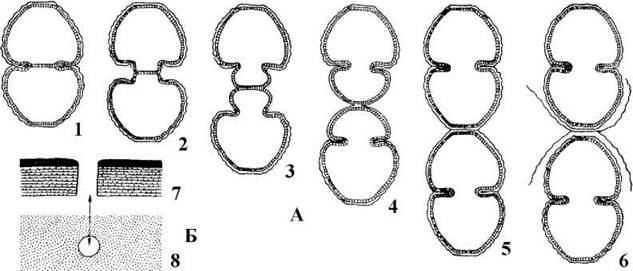
Рис. 24.72. Десмідіальні водорості. А - поділ клітини: 1 - утворення поперечної перегородки, 2 - подовження перешийка та початок розвитку молодих напівклітин, 3, 4 - поступовий ріст молодих напівклітин, 5, 6 - розділення сформованих напівклітин, що супроводжується скиданням їх тимчасової оболонки. Б - будова пори: 7 - на зрізі, 8 - при вигляді зверху (за Паламар-Мордвинцева, 1989).
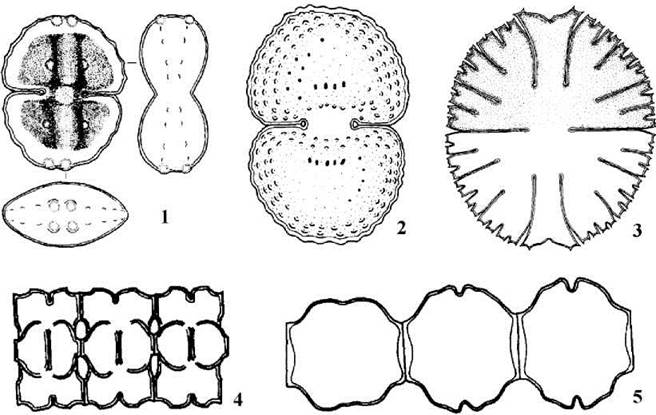
Рис. 24.73. Десмідіальні водорості: 1, 2 - деякі види роду Cosmarium (1 - C.bigemma; 2 - C.hornavense), 3 - Micrasterias, 4, 5 - деякі види роду Desmidium (4 - D.aptogonum; 5 - D.graciliceps) (Gontcharov, 1997; 1998).
Розмножуються десмідіальні вегетативним поділом або внаслідок кон’югації. При вегетативному поділі у перешийку за способом фрагмопласту утворюється клітинні перегородка. У кожної молодої клітини в зоні перешийку відбувається ріст, внаслідок чого будується друга напівклітина. Під оболонкою молодої напівклітини утворюється типова для десмідіальних водоростей тришарова оболонка, а зовнішній шар оболонки нової напівклітини скидається і клітини роз'єднуються (рис. 24.72).
При статевому розмноженні зигота (зигоспора) проростає звичайно двома проростками, оскільки з чотирьох ядер, що утворились внаслідок мейозу два ядра дегенерують.
Найбільш поширеними родами десмідіальних водоростей є Cosmarium та Micrasterias. Клітини Cosmarium поодинокі, глибоко перетягнуті, колоній не утворюють. Напівклітини більш-менш напівсферичні, і містять зірчасті хлоропласти (рис. 24.73).
Клітини Micrasterias сплощені, при вигляді зверху більш-менш округлі, глибоко перетягнуті. Напівклітини звичайно розділені на три великі лопаті - одну полярну та дві бічні. Кожна напівклітини містить один лопатевий хлоропласт.
У роду Desmidium клітини з'єднуються у стрічкоподібні колонії, звичайно більш-менш спірально скручені. Перетяжка, що поділяє клітину на напівклітини неглибока. Хлоропласти масивні, зірчасті, глибоколопатеві, по одному в кожній напівклітини.
Мешкають представники Desmidiales у прісних водоймах різного типу, проте найбільш числені вони у сфагнових болотах.