ФІЗІОЛОГІЯ ТА БІОХІМІЯ РОСЛИН
Опорний конспект лекцій
12. ФІЗІОЛОГІЯ РОЗМНОЖЕННЯ РОСЛИН
Вегетативне розмноження
Вегетативне розмноження - типова властивість рослин, яка відрізняє їх від тварин. Вегетативним розмноженням називається відтворення рослин з їх вегетативних частин, тобто з кусочків талому, пагона чи кореня.
Це - одна з форм безстатевого розмноження, дуже поширена серед рослин. Найскладніших та найрізноманітніших способів воно досягає у вищих і особливо - у квіткових рослин. Багато рослин розмножується тільки вегетативно. Це здійснюється як спеціалізованими, так і неспеціалізованими частинами тіла рослин. Під час вегетативного розмноження у багатьох рослин відбувається роз’єднання частин рослинного організму, які розвиваються в нові особини. Так, в елодеї можна відокремити частину тіла рослини, і вона розвиватиметься самостійно. Аналогічно розмножується ряска, яка за короткий час утворює величезну кількість рослин, що вкривають водну поверхню різних водойм.
Способи вегетативного розмноження рослин
Існують три основні способи вегетативного розмноження в природних умовах:
1) розмноження відсадками чи паростками різної природи, які відділяються від материнської особини після укорінення;
2) розмноження бульбами й цибулинами;
3) відособлення частин материнського організму.
Вегетативне розмноження рослин має величезне біологічне значення.
Здатність покритонасінних до інтенсивного вегетативного розмноження відіграла немалу роль у їх успішній конкуренції з голонасінними, яким властиве тільки насінне розмноження. У багатьох випадках вегетативне розмноження стало єдино можливою формою репродукції, яка забезпечує розселення виду й збереження генофонду. Досить часто рослини утворюють структури, спеціально призначені саме для вегетативного розмноження: цибулини, кореневища, столони, бульби. Деякі з них слугують для резервування поживних речовин, що дає змогу рослині пережити несприятливі умови. Тому більшість рослин зі спеціалізованими органами вегетативного розмноження практично втрачають здатність до розмноження насінням, передаючи функції як розмноження, так і відкладання та зберігання поживних речовин цим органам.
Між розвитком спеціалізованих органів вегетативного й статевого розмноження спостерігається декілька аналогій. І ті й інші утворюються в результаті індукції, яка сприймається листками і передається за допомогою хімічних стимулів відповідним частинам рослин. Усі головні етапи фотоперіодичної реакції при утворенні органів вегетативного розмноження також близькі до фотоперіодичних реакцій квітучих рослин. Після диференціації та росту в обох випадках настає етап дозрівання. При утворенні бульб та цибулин, як і при цвітінні, вегетативний ріст пригнічується. Морфологічно як квіти, так і бульби та цибулини є видозміненими пагонами.
Бульба являє собою потовщену здуту частину стебла, яка складається з декількох міжвузль.
Цибулина утворюється в результаті вкорочення міжвузль та іммобілізації вуглеводів в основі дуже молодих листків.
Розмноження бульбами. Здатність до утворення бульб формується в процесі онтогенезу поступово і проявляється після завершення ювенільного етапу. Бульби живуть лише один рік. У процесі бульбоутворення спочатку розвиваються столони - стебла зі зміненою геотропічною реакцією (перший етап), а потім на них формуються бульби (другий етап).
Найбільш суттєво на цей процес впливають температура й тривалість дня. При цьому частина рослин утворює бульби лише при короткому дні, а частина - як при короткому, так і при довгому. У дослідах із щепленням рослин, що не утворюють бульби, на бульбоноси (наприклад, томати на картоплю, соняшник на топінамбур) було показано, що стимул до утворення бульб у фотоперіодично чутливих бульбоносів не володіє видовою специфічністю, виробляється в листках і має гормональну природу.
З’ясувалося, що ауксини пригнічують бульбоутворення, гібереліни посилюють ріст столонів і, таким чином, сприяють росту бульб. А найбільший позитивний вплив на формування бульб мають цитокініни. У КДР перший етап контролюється співвідношенням АБК/ГА, а другий - ІОК/Цк. При довгому дні в листках утворюється більше гіберелінів, а в стеблових бруньках - ауксину. В цих умовах проходить перший етап і формуються столони. При короткому дні в коренях і в столонах є багато цитокінінів, а в листках - абсцизової кислоти. Настає другий етап, суть якого полягає в гальмуванні утворення столонів та запуску процесів бульбоутворення.
На думку М.Чайлахяна (1984), механізм гормональної індукції бульбоутворення (перехід до другого етапу і його реалізація) у видів, які утворюють бульби при короткому дні, складається з двох фаз. Спочатку з листків до нижніх бруньок надходять абсцизини і гібереліни. Оскільки при КД співвідношення їх (АБК/ГА) зміщене із переважанням абсцизинів, то прискорений ріст столонів, викликаний гібереліном, гальмується (перша фаза).
Під час другої фази утворюються бульби, ріст яких регулюється більш високою концентрацією цитокініну в коренях і столонах відносно ауксинів (співвідношення ІОК/ЦК зменшується).
Таким чином, в умовах ДД при несприятливому для бульбоутворення співвідношенні гормонів (у листках переважає ГА, в коренях - ІОК) відбувається утворення й ріст коренів та столонів. Стимулом для бульбоутворення служить таке співвідношення АБК до ГА, яке впливає на завершення першого етапу та переходу до першої фази другого етапу.
Цим різноманітність способів бульбоутворення у рослинному світі не обмежується. Так, бульби бувають стеблового і кореневого походження, підземні і надземні. У жоржини бульби кореневого походження, формуються на коренях і стеблові пагони при цьому не утворюються. Бульбами розмножуються картопля, топінамбур, батат та інші рослини.
Розвиток та ріст бульб підтримується надходженням до них продуктів фотосинтезу, які утворюються в листках. При цьому інтенсивність транспорту асимілятів до бульб корелює з вмістом у них цитокінінів, ауксину та гіберелінів, які створюють атрагуючу силу бульб. Власне тому при інтенсивному утворенні бульб затримується ріст надземних частин рослин.
Розмноження цибулинами. Розмноження цибулинами характерне для однодольних рослин з родин лілійних та амарилісових: тюльпана, нарциса, гіацинта, лілій тощо. Дикий часник утворює велику кількість цибулинок: на орному гектарі їх може бути до 600 кг.
Цибулина складається із дуже короткого стебла та м’ясистих листків із резервними речовинами. Зверху вона вкрита залишками минулорічних листків, запасні речовини яких уже втрачені. Цибулина містить одну або кілька дочірніх цибулин («діток», зубків), кожна з яких може утворювати пагін. Під кінець вегетації кожен пагін дає нову цибулину. Корені у цибулин відходять безпосередньо від стебла, і головного кореня немає.
Якщо більшість бульбоносів утворюють бульби в умовах короткого дня, то формування цибулин, навпаки, вимагає довгого дня. В дослідах витримування навіть частини листків при короткому дні гальмувало формування цибулин. Регуляція цього процесу вивчена недостатньо. Органами, що рецептують дію фотоперіоду, є листки.
Фотоперіодичний стимул може передаватися з одного пагона на інший тільки за умови, що у другого пагона видалені листки. Коли пагони з листками піддаються дії різних фотоперіодів, то цибулину формує лише пагін, який знаходився на ДД. Стимулом до утворення цибулини є певне співвідношення фітогормонів, або комплекс фітогормонів. Так, формування цибулин супроводжується зменшенням активності ауксинів, а збільшення їх концентрації прискорює ріст листків і цибулинних лусок.
Вегетативне розмноження цибулинних рослин не так пов’язано з формуванням цибулини, як з утворенням на ній пазушних пагонів - майбутніх дочірніх цибулинок. Вони закладаються в період зимового спокою. В цей час, хоч і дуже повільно, формуються нові органи. При проходженні періоду спокою у цибулинних рослин, на відміну від бруньок і насіння, ростові процеси повністю не зупиняються. Кінетин, гіберелова та індолілоцтова кислоти посилюють утворення пазушних бруньок, які формують дочірні цибулини. Такий самий ефект дають підвищення температури зимового зберігання цибулин та видалення квітконосного пагона.
Проростання цибулин, як і бульб, відбувається після проходження періоду спокою. Вихід із стану спокою прискорюється після дії на цибулини низької температури, тобто яровизацією. Стан спокою цибулин підтримується високим вмістом АБК, а при проростанні рівень інгібіторів знижується, а цитокінінів, ауксинів і гіберелінів - зростає.
Знання принципів регуляції стану спокою бульб і цибулин дозволяє запобігати їх проростанню при зберіганні з допомогою синтетичних інгібіторів росту, таких, як гідразид малеїнової кислоти.
Розмноження іншими способами
У дикоростучих рослин широко поширене розмноження кореневою поростю, тобто відростанням при пошкодженні рослин сплячих бруньок у нижній частині стовбура (пенькові пагони). Таке розмноження характерне для вишні, сливи, яблуні, айви, бузку, глоду, вільхи, берези, хріну, люцерни, осоту та інших рослин. Часто на пошкоджених коренях коло поверхні землі виникають кореневі пагони, які розвиваються з адвентивних бруньок.
Деякі види розмножуються кореневищами (пирій повзучий, м’ята перцева та ін.). Кореневища є підземними стеблами, які ростуть горизонтально і мають дрібні плівчасті бурувато-коричневі листочки, бруньки, додаткові корені.
Кореневища є і зимуючим органом, і органом розмноження.
Трапляється також розмноження надземними пагонами, наприклад, столонами (зеленчук, костяниця, перстачі та ін.) чи вусами (суниці, полуниці, жовтець тощо). Столони - це повзучі горизонтальні стебла, які стеляться по поверхні землі. Вони не є зимуючим органом. Від їх вузлів відходять додаткові корені, які разом із частиною стебла відокремлюються від материнського куща, і таким чином утворюються нові рослини.
Надземні повзучі пагони (вуса, столони, батаги) є переходом від типових вертикальних стебел до кореневища.
Деякі рослини розмножуються гілками-відсадками, що вкорінюються на місці контакту пагона з ґрунтом.
Є група живородних рослин. У пазухах їх листків та в суцвіттях замість квіток утворюються маленькі, вкриті листками пагони, які, опадаючи з материнської рослини, вкорінюються. Живородні рослини поширені переважно в полярних, високогірних і степових місцевостях.
Деякі способи вегетативного розмноження тісно пов’язані з процесами регенерації (живцювання, вівіпарія й ін.). Так, утворення вивідкових бруньок (вівіпарія) може розглядатися як випадок соматичного ембріогенезу (каланхое) і є результатом фізіологічної диференціації тканин. Різноякісність тканин пов’язана з реакцією на довгий день і полягає в збільшенні на краях листка вмісту хлорофілу, ДНК, РНК, білків. Таким чином, утворення вивідкових бруньок, як спеціалізованого органу вегетативного розмноження контролюється фотоперіодом.
Використання вегетативного розмноження в рослинництві
У рослинництві найчастіше застосовують розмноження живцями, щепленням та відсадками.
Відсадками - шляхом пригинання нижніх галузок і присипання їх землею. Укорінений пагін морфологічно не відрізняється від інших.
Живцями. Використовують зимові безлисті та літні з листками живці, зелені листки (бегонія). Зимові живці беруть з однорічних, дво-, трирічних гілок. Найбільш придатні молодші, однорічні живці після листопаду 30 см завдовжки. Зелені живці зрізають тоді, коли в стеблі відбувається приріст і вони остаточно не здерев’яніли. На поверхні зрізу утворюється плівка, під якою енергійно діляться клітини і пізніше утворюється калус. На калусі розвиваються корені, а пагони утворюються з бруньок живця.
Живці, взяті з різних ділянок рослин, мають різні властивості. Живці з плодоносної зони дають рослини, які швидше зацвітають порівняно з рослинами з ювенільних живців. Живці з бокової зони часто дають рослини, які довго зберігають двосторонню будову. Живці зі старих дерев укорінюються гірше. Має значення і фізіологічний стан самого живця. Максимальною здатністю до вкорінення володіють пагони в період сповільнення їх вегетативного росту, коли вміст у них ауксинів найбільший.
Обробляючи живці різними ауксинами (ІОК, індолілмасляна, індолілпропіонова, трийодбензойна, а-нафтилоцтова кислоти), стимулюють коренеутворення у видів, які самі не вкорінюються.
Розвиток живця значно прискорюється завдяки щепленню й використанню кореневої системи підщепи. При цьому прищепа зберігає свої властивості. Рослину, на якій роблять щеплення, називають підщепою. Підщепу підбирають так, щоб вона мала певні господарськоцінні якості - морозостійкість, сильний або карликовий ріст, довговічність, стійкість до хвороб тощо.
Є різні види щеплення (Проценко, 1978) - щеплення під кору, щеплення врозщеп, окулірування (щеплення однією брунькою-вічком), копулірування, коли прищепа та підщепа мають однакову товщину.
Щеплення під кору у деревних порід проводять навесні, коли у рослин спостерігається велика активність камбію. Кора в цей час легко відокремлюється від деревини стебла. На підщепі роблять горизонтальний зріз на міжвузлі під стебловим вузлом. Потім кору надрізають у вертикальному напрямку вниз від зрізу й обережно відгинають край кори. Прищепу беруть у вигляді живця з трьома-чотирма міжвузлями. На нижньому кінці міжвузля прищепи відрізають напівконус у вигляді язичка, який вставляють під кору підщепи так, щоб опукла сторона язичка була назовні. При цьому відігнуті кінці кори добре притискують і місце щеплення обв'язують плівкою.
Щеплення врозщеп застосовують у випадках, коли підщепа значно товща, ніж прищепа. При цьому підщепу розколюють і в щілину вставляють живці прищепи, кінці яких загострюють клином. Місце щеплення замазують садовим варом і теж обв'язують. Цим способом щеплять як дерев'янисті, так і трав'янисті рослини.
Дуже часто застосовують окулірування — щеплення однією брунькою (вічком). При цьому бруньку вирізають із кусочком кори та деревини і прищеплюють на дичках. На підщепі роблять Т-подібний надріз, а вічко вставляють із так званим щитком, тобто з кусочком деревини та кори. Після того, як щиток уставлено під кору, його притискують корою та обв'язують. Якщо прищепа та підщепа мають однакову товщину, застосовують копулірування. Для цього роблять навскісні зрізи на прищепі та підщепі, щоб вони щільно прилягали одна до одної. Такі навскісні зрізи роблять для того, щоб збільшити поверхню стикання прищепи та підщепи. Місце щеплення обв'язують і змащують садовим варом. Щеплення проводять навесні та восени, але воно можливе і в іншу пору року, що залежить від способу щеплення. Так, часто окулірують влітку сплячою брунькою, яка до початку зими зростається з підщепою і навесні наступного року починає ріст.
Зростання прищепи та підщепи відбувається таким чином. Живі клітини коло поверхні зрізу, особливо клітини камбію, кори, судинних пучків, починають інтенсивно ділитися, заповнюють щілину проміжною тканиною. Через деякий час клітини прищепи та підщепи з’єднуються плазмодесмами, потім утворюються диференційовані клітини, які з’єднують провідні системи підщепи та прищепи.
Загалом методи вегетативного розмноження набули широкого поширення в рослинництві. В останній час у практиці сільського господарства та селекційній роботі широко використовують метод отримання рослин із потрібними властивостями з допомогою культури тканин (а також меристем, сім’ядолей, зародків).
Із однієї клітини чи з групи клітин у стерильних умовах вирощують цілі рослини для клонального розмноження цінних культур, отримують безвірусні форми сільськогосподарських рослин (картопля, цукровий буряк, виноград), шляхом злиття протопластів створюють міжвидові гібриди; культивуванням мікроспор отримують дигаплоїдні рослини, дуже цінні для селекції; розвиваються методи культивування частин гаметофіту і репродуктивних органів. Цей метод широко застосовується для створення банку безвірусних сортів багатьох культур.
Розробка основ методу має порівняно недавню історію. Німецький ботанік Габерландт (1902) вперше застосував цей метод для роботи з клітинами палісадної паренхіми. Він хотів виростити цю тканину з однієї клітини. Невдача, що спіткала автора, дещо затримала роботу з рослинними організмами, але дослідження були продовжені на тваринних об’єктах.
В Україні культуру ізольованих коренів широко використовував М.Г. Холодний ще в 1915 р., коли він за допомогою даного методу виявив біосинтез фітогормонів у кінчиках коренів, що стало основою для розробки його теорії тропізмів. Цей метод він використовував і пізніше.
В 30-і роки праці американського вченого Уайта та французького вченого Готре зумовили розробку власне сучасного методу культури рослинних тканин і органів. Уайт довів, що коренева меристема може рости тривалий час, якщо її пересаджувати на свіже середовище. Культура клонів кінчиків кореня помідорів в його дослідах підтримувалася понад 30 років (Уайт, 1949).
Найважливішою умовою вирощування ізольованих рослинних тканин є поживні середовища необхідного складу. Заслугою Готре є те, що він вдосконалив склад поживних середовищ для вирощування недиференційованих тканин різних органів однодольних, дводольних та голонасінних рослин (1959). Готре ввів у культуру різні калусні тканини у вигляді чистих клонів. В Україні ці роботи були організовані в лабораторії Ф.Л. Калініна (Калінін, Сарнацька, Поліщук, 1980).
Відомо, що клітини різних тканин, органів, організмів функціонують in vivo в специфічних метаболічних умовах. Це є науковою основою для розробки різних за складом поживних середовищ. Як відомо, тепер їх створено більше ста. Обов'язковими компонентами поживних середовищ є певні макро- і мікроелементи, вуглеводи, біологічно активні речовини, які регулюють процеси поділу клітин та їх диференціацію. Склад найбільш вживаних в технології мікроклонального розмноження середовищ висвітлено в монографіях, статтях, наукових розробках. Мінеральний компонент, обумовлений основними фізіологічними вимогами більшості тканин рослин, є найбільш стабільним. Модифікації вмісту поживних речовин досягаються внесенням різноманітних регуляторів росту. Особливо важлива регуляторна роль ауксинів і цитокінінів та їхнього співвідношення.
Метод культури ізольованих тканин полягає ось у чому. Шматочки тканин вміщують у пробірку, в колбочки, чашки Петрі на поживні рідини або напівтверді агарові середовища. Окремі тканини вирощують у стерильних умовах. Таким способом ізольовану тканину можна витримувати протягом десятків років у стані активного росту. У процесі культури ізольованих тканин перш за все утворюється калюс, у ньому закладаються бруньки, з яких розвиваються пагони, а також додаткові корені.
Калюс, калус - маса недиференційованих клітин, яка утворюється в разі пошкодження рослин, виникає внаслідок розростання групи недиференційованих клітин, утворюваних камбієм чи іншими тканинами в місцях надрізів, тріщин, ран; розвивається під час культивування на штучних середовищах з поодинокої клітини, має вигляд аморфної маси, тонкого шару або напливу.
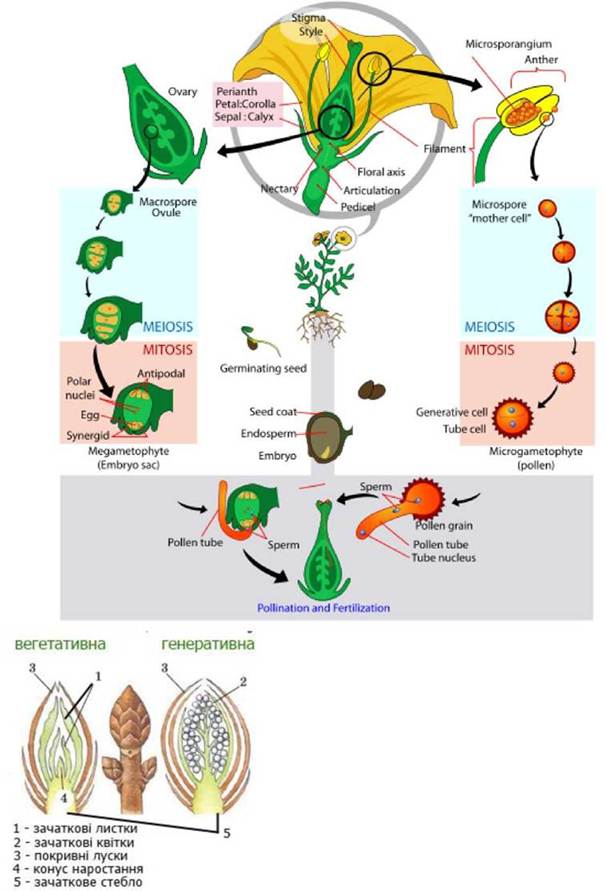
Рис. 36. Будова бруньки.
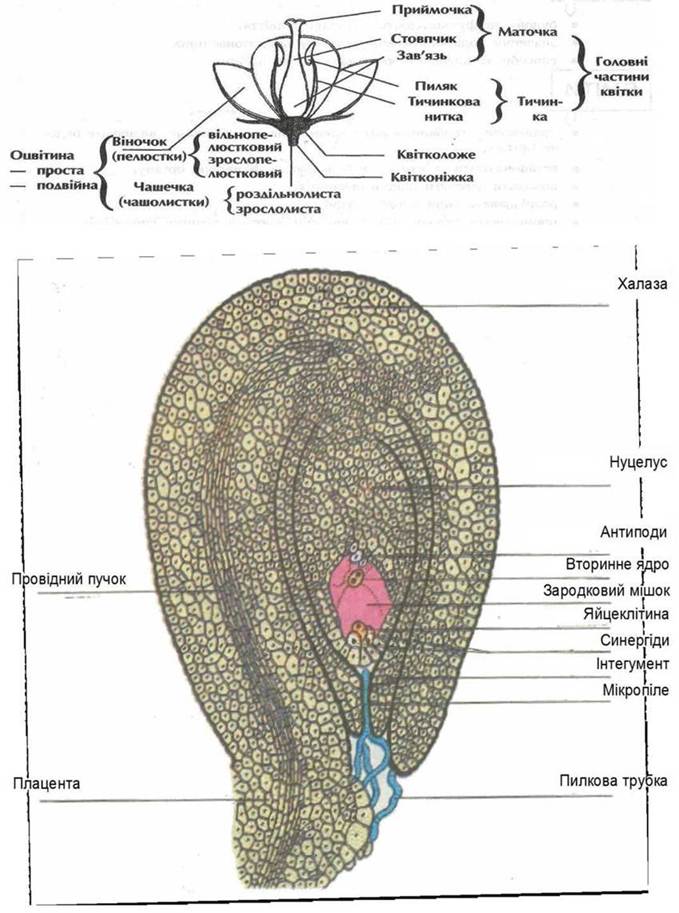
Рис. 37. Будова квітки.
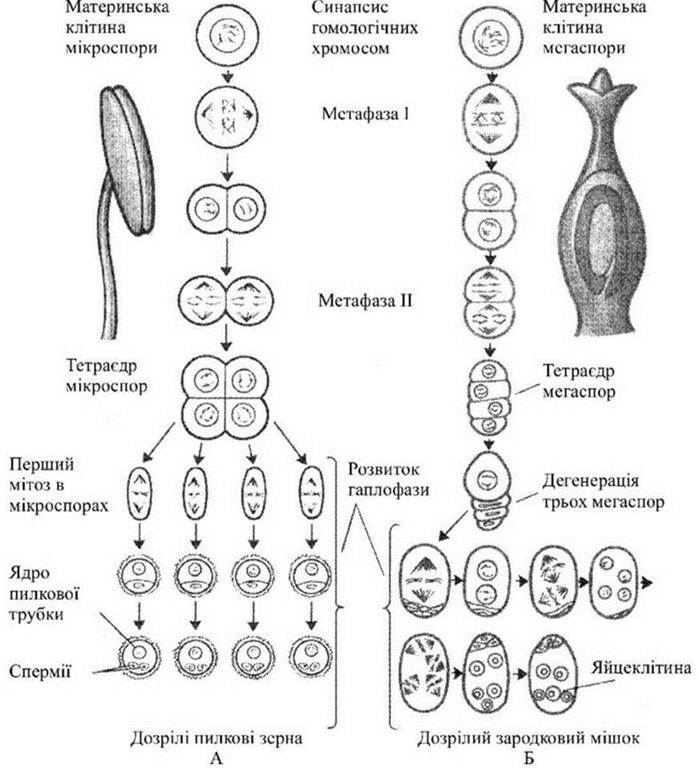
Рис. 38. Будова насіннєвої бруньки ірису.
Спорогенез та гаметогенез у квіткових рослин
Спорогенез починається з мейозу диплоїдних спорогенних клітин пилкового мішка та археспоріальної клітини нуцелуса і завершується утворенням мікро- та мегаспор.
Гаметогенез відбувається шляхом мітозу. З кожної мікроспори у результаті двох мітотичних поділів формується гаплоїдний чоловічий гаметофіт з двома сперміями та одним вегетативним ядром. Гаплоїдне ядро єдиної мегаспори ділиться мітозом чотири рази і з утворених восьми ядер формується семиклітинний зародковий мішок (жіночий гаметофіт).
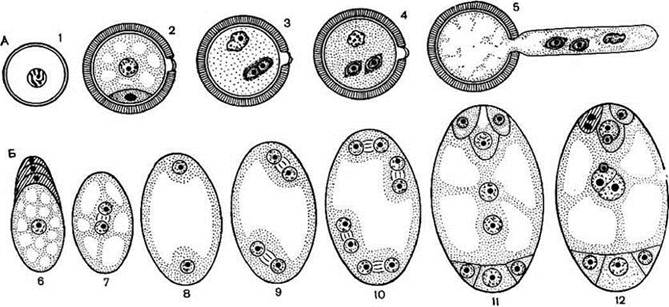
Рис. 39. Гаметогенез: розвиток пилкового зерна (А) і зародкового мішка (Б).
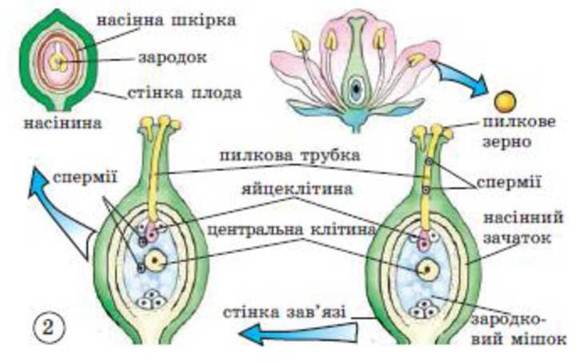
Рис. 40. Подвійне запліднення.
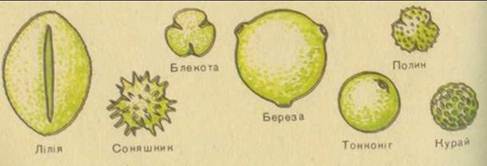
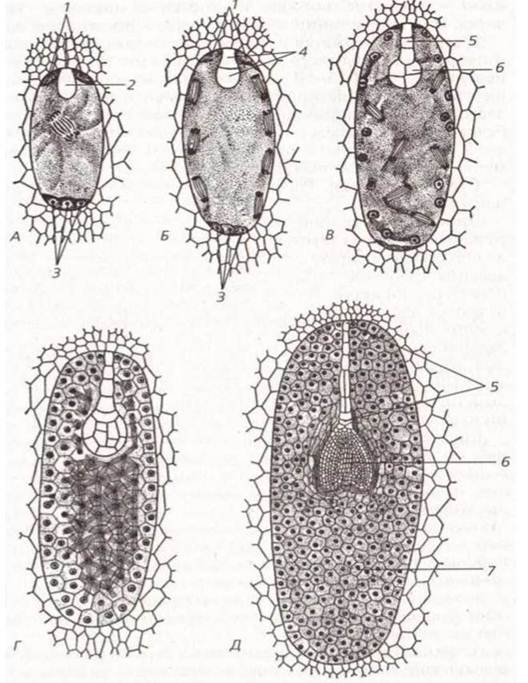
Рис. 41. Форми пилкових зерен.
Розвиток зародка і ендосперму у дводольних: А - стан спокою зиготи, початок ділення вторинного ядра і формування ендосперму; Б, В - зародок у фазі проембріо, швидкий мітотичний поділ ядер майбутніх клітин ендосперму; Г - глобулярна фаза розвитку зародка, формування клітин ендосперму, які продовжують ділитися; Д - серцевидна фаза розвитку зародка, сформований ендосперм; 1 - синергіди, 2 - зигота, 3 - антиподи, 4 - двоклітинний зародок, 5 - суспензор, 6 - зародок, 7 — ендосперм.
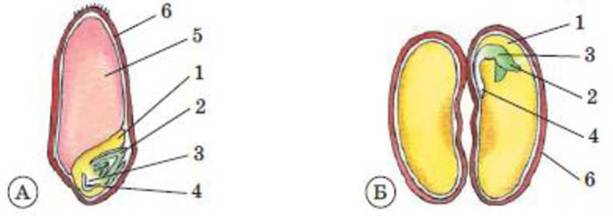
Рис. 42. Будова насінини пшениці (А) та квасолі (Б): 1 - сім’ядоля (у насінини квасолі дві сім'ядолі); 2 - зародкова брунька; 3 - зародкове стебло; 4 - зародковий корінь; 5 - ендосперм; 6 - насінна шкірка (у насінині пшениці вона зрослася зі стінкою плода).
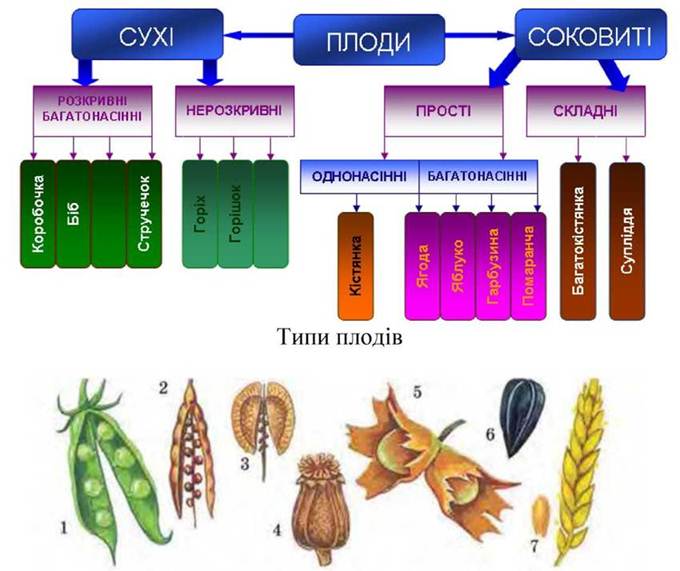
Рис. 43. Сухі плоди: 1 - біб; 2 - струнок; 3 - стручечок; 4 — коробочка; 5 - горіх; 6 - сім'янка, 7 — зернівка.
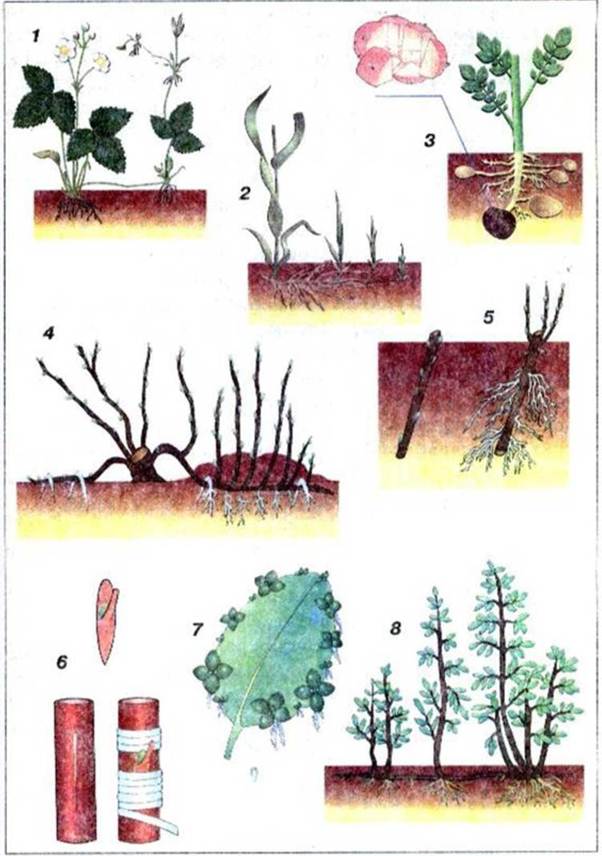
Рис. 44. Вегетативне розмноження:
1 - вусами; 2 - кореневищами; 3 - бульбами; 4 - підводками; 5 - стебловими живцями; 6 - щепленням; 7 - виводковими бруньками; 8 - кореневими живцями.
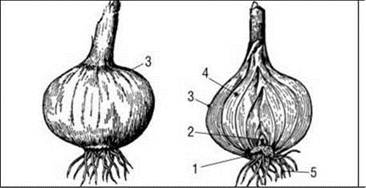
Рис. 45. Будова цибулини:
а — проста плівчаста цибулина зовні і в розрізі: 1 — укорочене стебло - денце; 2 — брунька; 3 — сухі плівчасті луски; 4 — соковиті луски; 5 — додаткові корені.
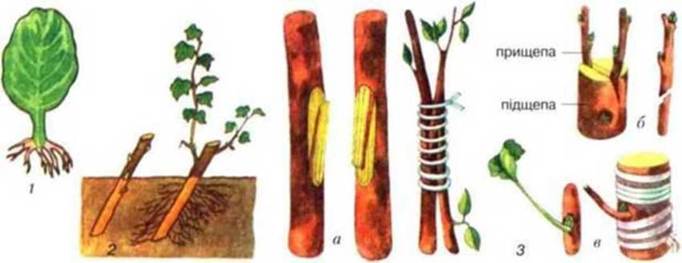
Рис. 46. Способи вегетативного розмноження:
1 — розмноження листком (сенполія): 2 — живцювання; З — щеплення (а — зближенням. б— живцем, в — вічком).

